|
|
|
Лекція №2
БІОЛОГІЧНІ МЕМБРАНИ. МЕМБРАННИЙ ТРАНСПОРТ.
План
1. Структурні елементи біомембран. Фізичні властивості. Рідкокристалічний стан мембран. Динамічні властивості.
2. Пасивний транспорт речовин крізь мембранні структури. Рівняння Фіка. Коефіцієнт проникності мембрани для певної речовини. Рівняння Нерста-Планка.
3. Активний транспорт, основні види.
4. Спряження потоків. Швидкість дифузії.
5. Мембранні потенціали спокою.
6. Потенціал дії (ПД). Еквівалентна електрична схема мембрани. Феноменологічні рівняння Ходжкіна—Хакслі. Поняття про воротні йонні струми. Рівняння Ходжкіна—Хакслі для процесу поширення ПД в нервових волокнах. Швидкість і особливості поширення ПД в аксонах.
1. Структурні елементи біомембран. Фізичні властивості. Рідкокристалічний стан мембран. Динамічні властивості.
Важливою частиною клітини являються біологічні мембрани. Вони відмежовують клітину від оточуючого середовища, захищають її від шкідливих зовнішніх дій, керують обміном речовин між клітиною та її оточенням, сприяють генерації електричних потенціалів, беруть участь в синтезі універсальних акумуляторів енергії АТФ в мітохондріях ті інше. Мембрани формують структуру клітини забеспечуючи її функції.
Мембрани оточують всі клітини, внутрішньоклітинні мембрани розділяють клітину на ряд закритих відсіків (компартаментів), кожний з яких виконує визначену функцію. Товщина мембрани декілька нанометрів і тому її побачити можливо тільки за допомогою електронного мікроскопу.
Вивчення будови мембрани та її властивостей стало можливим при використанні фізико-хімічної моделі мембрани (штучної мембрани). Найбільшого розповсюдження отримали три моделі:
-
перша модель – моно шари фосфоліпідів на межі поділу вода-повітря або вода-масло;
-
друга модель – ліпосоми – мілкі кульки (везикули), які складаються з біліпідної мембрани й отримані при обробці суміші води та фосфоліпідів ультразвуком;
-
третя модель, яка дозволила вивчати деякі властивості біомембран прямими методами, була біліпідна (бішарова ліпідна) мембрана.
Фізичні властивості та параметри мембран. Фосфоліпіди в мембрані, за умов її нормального функціонування, знаходяться в рідкокристалічному стані. Рідинні якості мембрани підтверджуються ЕПР і ЯМР. В’язкість ліпідного шару мембрани приблизно на два порядки вище в’язкості води, вона дорівнює 30-100 мПа*с, що приблизно дорівнює в’язкості рослинного масла. Поверхневий натяг на 2-3 порядки нижче ніж у води. При зміні температури в мембрані можна спостерігати фазові перетворення: плавлення ліпідів при нагріванні та кристалізацію при охолодженні. Подвійний шар фосфоліпідів робить мембрану клітини подібною до ємності, електроємність 1 мм2 мембрани складає 5-13 нФ.
2. Пасивний транспорт речовин крізь мембранні структури. Рівняння Фіка. Коефіцієнт проникності мембрани для певної речовини.
Рівняння Нерста-Планка.
Важливим елементом функціонування мембран являється їх спроможність пропускати або не пропускати молекули (атоми) та іони. Як синонім переносу частинок в біофізиці широкого розповсюдження набув такий термін як транспорт частинок.
Пасивний транспорт – транспорт речовин за градієнтом концентрації, що не потребує витрат енергії. Пасивно відбувається транспорт гідрофобних речовин крізь ліпідний бішар. Пасивний транспорт, в якому беруть участь мембранні білки, називають полегшеною дифузією.
Закони Фіка. В системі з градієнтом концентрації речовини dC/dx в напрямку х дифузійний потік J визначається першим законом Фіка:
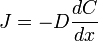 , ,
де D – коефіцієнт дифузії (знак "-" вказує на напрямок потоку від більших концентрацій до менших).
У разі градієнту концентрації не лише в напрямку х, треба використовувати загальнішу формулу:
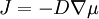 , ,
де μ - хімічний потенціал.
В загальному випадку перенос іонів визначається двома факторами: нерівномірністю їх розподілення, тобто градієнтом концентрації, та дією електричного поля:
J = -D(dc/dx)- umZFс(dφ/dx) - рівняння Нернста-Планка.
Електрохімічний потенціал – термодинамічна функція, що характеризує стан якого-небудь компоненту, що складається із заряджених частинок (електронів, іонів), у фазі даного складу. Електрохімічний потенціал може бути визначений як приріст будь-якого з термодинамічних потенціалів системи при введенні в неї однієї зарядженої частинки i-того компоненту при незмінних решти всіх змінних, від яких залежить даний потенціал. Таким чином, різність електрохімічних потенціалів між двома точками є енергією, яка буде виділена в системі при переміщенні частинки між ними. Електрохімічний потенціал виражається формулою:
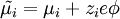 , ,
де 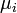 – хімічний потенціал i-того компоненту, – хімічний потенціал i-того компоненту, 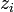 – заряд частинки, – заряд частинки, 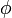 – електричний потенціал, е – елементарний заряд; останній доданок виражає роботу по подоланню електричних сил. Якщо електрохімічний потенціал відноситься до 1 молю речовини, цей член дорівнює – електричний потенціал, е – елементарний заряд; останній доданок виражає роботу по подоланню електричних сил. Якщо електрохімічний потенціал відноситься до 1 молю речовини, цей член дорівнює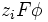 , де F– число Фарадея. , де F– число Фарадея.
3. Активний транспорт, основні види.
Активний транспорт – опосередкований транспорт біомолекул, неорганічних іонів та малих молекул через цитоплазматичну або будь-яку іншу мембрану клітини. На відміну від пасивного транспорту, цей процес вимагає витрат хімічної енергії у формі АТФ або різності концентрацій іншої речовини з двох боків мембрани. У цій формі транспорту, молекули рухаються проти або електричного потенциалу, або концентраційного градієнту (колективно названими електрохімічним потенціалом).
Відомі два головні типи активного транспорту: первинний і вторинний.
У первинному транспорті, енергія АТФ безпосередньо використовується для перенесення бажаної молекули або іону через мембрану незалежно від будь-яких інших молекул. Більшість ферментів, які використовують цей вид транспорту, — трансмембранні АТФази. Найпоширенішою АТФазою, універсальною для всіх форм клітинного життя, є калій-натрієвий насос, який допомогає підтримувати клітинний потенціал.
Вторинний транспорт вимагає використання дифузії іншої молекули через мембрани для того, щоб перенести бажану молекулу. У відтворенні механізму вторинного активного транспорту АТФ не використовується, натомість, енергія береться за рахунок різниці електрохімічних потенціалів інших молекул.
Існує три форми вторинного активного трансторту: антипорт, сімпорт, уніпорт.
Антипорт. Під час антипорту, два різних іона або молекули проходять мембрану у протилежний напрямках. Одна з цих полекул рухається уздовж градієнту електрохімічного потенціалу, звільняючи енергію, яка, натомість, використовується для перенесення іншої молекули. Прикладом є натрій-кальцієвий антипортер, який дозволяє трьом натрієвим іонам проходити до клітини в обмін на вихід одного іону кальцію, який входить до клітини.
Багато клітин також мають кальцієву АТФазу, яка може діяти при нижчих внутрішньоклітинних концентраціях кальцію і встановлює нормальний стан концентрації цього важливого іону. Але АТФаза експортує іони кальцію повільніше: тільки 30 іонів за секунду проти 2000 за секунду у антипортера. Антипортер вступає в гру, коли концентрація кальцію різко підвищується і гарантує швидке оновлення. Це показує, що єдиний вид іону може бути перенесений кількома ферментами, яким не потрібно бути активними весь цей час.
Симпорт. Симпорт також використовує потік інших іонів або молекіл уздовж електрохімічного градієнту для перенесення іншої молекули проти, але дві солекули рухаються у одному напрямку. Приклад – глюкозний симпортер, який переносить два іони натрію разом з молекулою глюкози, яку він імпортує в клітину.
Уніпорт – транспорт однієї речовини в одному напрямку в залежності від градієнта. Уніпорт здійснює, наприклад, потенціал-залежний натрієвий канал, крізь який в клітину під час генерації потенціалу дії переміщуються іони натрію.
Молекулярна організація активного транспорту на прикладі роботи Na+ - К+ насосу. В першу чергу цей переносник приєднує з внутрішньої сторони мембрани три іони Na+. Ці іони змінюють конформацію активного центру Ат фази. Після такої активації Ат фаза здатна гідролізувати одну молекулу АТФ, причому фосфат-іон фіксується на поверхні переносника з внутрішньої сторони мембрани.
Енергія, що виділяється, витрачається на зміну конформації АТФази, після чого три іони Na+ та іон РО43- (фосфат) опиняються на зовнішній стороні мембрани. Тут іони Na+ від’єднуються, а РО43- заміщується двома іонами К+. Потім конформація переносника змінюється на початкову, а іони К+ опиняються на внутрішній стороні мембрани. Тут іони К+ від’єднуються, переносник знову готов до роботи.
Більш коротко дію АТФази можна зобразити так:
-
вона з середини клітини “забирає” три іони Na+, потім розщеплює молекулу АТФ та приєднує до себе фосфат;
-
“викидає” іони Na+ та приєднує два іони К+ із зовнішнього середовища;
-
від’єднує фосфат, два іони К+ залишає в середині клітини.
В результаті у позаклітинному середовищі створюється висока концентрація іонів Na+, а всередині клітини – висока концентрація К+. Робота Na+, К+- АТФаза створює не тільки різницю концентрацій, а й різницю зарядів (вона працює як електрогений насос). На зовнішній стороні мембрани створюється додатний заряд, на внутрішній – від’ємний.
4. Спряження потоків. Швидкість дифузії.
Мембранні потенціали спокою.
Потенціалом спокою (ПC) називають мембранний потенціал збудливої клітини в стані спокою.
Природа мембранного потенціалу спокою. Для того, щоб на мембрані підтримувалася різниця потенціалів необхідний певний зміст різних іонів всередині і зовні клітини.
Згодом було доведено, що основний внесок у створення потенціалу спокою вносить калієвий струм, який виходить через специфічні білки-канали - калієві канали постійного струму. У спокої калієві канали відкриті, а натрієві канали закриті. Іони калію виходять з клітини за градієнтом концентрації, що створює на зовнішній стороні мембрани надлишок позитивних зарядів; на внутрішній стороні мембрани створюється негативний заряд. Деякий (невеликий) внесок у створення потенціалу спокою вносить також робота натрій-калієвої АТФази.
Потенціал спокою для більшості нейронів становить величину порядку -60 мВ…-70 мВ. У клітин невозбудімих тканин на мембрані також є різниця потенціалів, різна для клітин різних тканин і організмів.
Мембрани нервових і м'язових клітин (волокон) здатні змінювати іонну проникність у відповідь на зрушення мембранного потенціалу. При збільшенні ПП (гіперполяризацію мембрани) проникність поверхневих клітинних мембран для Na + і К + падає, а при зменшенні ПП (деполяризація) вона зростає, причому швидкість змін проникності для Na + значно перевищує швидкість збільшення проникності мембрани для К +.
5. Потенціал дії (ПД).
Потенціал дії (ПД) – це короткочасні високо амплітудні зміни мембранного потенціалу спокою (МПС), що виникають при збудженні живої клітини. По суті це електричний розряд – швидка короткочасна зміна потенціалу на невеликій ділянці мембрани збудливості клітини (нейрона або м'язового волокна), в результаті якого зовнішня поверхня цієї ділянки стає негативно зарядженою по відношенню до сусідніх ділянок мембрани, тоді як його внутрішня поверхня стає позитивно зарядженою по відношенню до сусідніх ділянок мембрани. Потенціал дії є фізичною основою нервового або м'язового імпульсу, який відіграє сигнальну (регуляторну) роль. Потенціал дії - один з найважливіших компонентів процесу збудження. У нервових волокнах він забезпечує проведення збудження від чутливих закінчень (рецепторів) до тіла нервової клітини і від неї - до синаптичних закінченням, розташованим на різних нервових, м'язових або залізистих клітинах. Вступаючи до ефекторні закінчення, ПД викликає виділення (секрецію) певної порції специфічних хімічних речовин, т. зв. медіаторів, що надають збудливу або гальмівний вплив на відповідні клітини.
Потенціали дії можуть відрізнятися за своїми параметрами в залежності від типу клітини і навіть на різних ділянках мембрани однієї і тієї ж клітини. Найхарактерніший приклад відмінностей: потенціал дії серцевого м'яза і потенціал дії більшості нейронів. В основі будь-якого потенціалу дії лежать наступні явища:
-
мембрана живої клітини поляризована – її внутрішня поверхня заряджена негативно по відношенню до зовнішньої завдяки тому, що в розчині біля її зовнішньої поверхні знаходиться більша кількість позитивно заряджених частинок (катіонів), а біля внутрішньої поверхні – більша кількість негативно заряджених часток (аніонів);
-
мембрана має вибіркову проникність – її проникність щодо різних частинок (атомів або молекул) залежить від їх розмірів, електричного заряду і хімічних властивостей;
-
мембрана збудливої клітини здатна швидко змінювати свою проникність для певного виду катіонів, викликаючи перехід позитивного заряду з зовнішньої сторони на внутрішню.
Перші дві властивості характерні для всіх живих клітин. Третя ж є особливістю клітин збудливих тканин і причиною, за якою їх мембрани здатні генерувати і проводити потенціали дії.
Основною математичною моделлю, що описує генерацію та передачу потенціалу дії, є модель Ходжкіна-Хакслі – математична модель, яка описує генерацію та розповсюдження потенціалів дії в нейронах та інших електрично збудливих клітинах – таких, наприклад, як серцеві міоцити. Модель являє собою комплекс ординарних диференційних рівнянь, котра змальовує характеристики електричного сигналу.
Електричний струм, що проходить через іонні канали, може бути математично виражений наступним рівнянням:
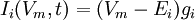
,
де Еі – рівноважний потенціал і-го іонного каналу. У випадку потенціал-залежних іонних каналів канальна провідність gі є функцією часу та потенціалу (електричної напруги), в той час як пасивна провідність є величиною сталою. Струм, генерований іонними транспортерами, залежить від виду іонів, що його переносить відповідний транспортер.
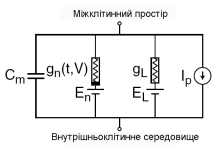
Подвійнному ліпідному шару клітинної мембрани відповідає електроємність Сm (рис. 3.1). Потенціал-залежні іонні канали відповідають нелінійній електричній провідності (gn, де n – окремий вид іонних каналів); це означає, що проводимість є потенціал- та час-залежною величиною. Канали мембранних пор відповідають пасивній провідності gL. Електрохімічний градієнт, що спонукає іони до руху крізь мембранні канали, показаний за допомогою акумуляторів з відповідною електрорушійною силою En та EL. Іонні транспортери відповідають джерелам струму Ip.
|
|


