Презентація: "Позахромосомне успадкування ознак"


Позахромосомне успадкування

Позахромосомне успадкування Позахромосомним успадконуванням називають процес передачі нащадкам спадкових детермшантів не хромосомными структурами клітини. Сукупність генів, що знаходяться в цитоплазмі еукариотичної клітини, називають плазмоном або плазмотипом. А гени, що входять до складу плазомну - плазмогени, або позаядарні гени. Ці елементи знаходяться у цитоплазматичних структурах: пластиди, мітохондрії, внуришньоклітинні паразити та ендосімбіонти, а у прокаріот - плазміди. Найважливішими носіями генетичної інформації в цитоплазмі еукариотів є геноми мітохондрій і пластид. Гени ДНК мітохондрій складають т.з. хондріом, а гени ДНК пластид – пластидом або пластом. Всі ці гени передаються нащадкам разом з цитоплазмою статевих клітин і незалежно від генів ядра.

Цитоплазматична спадковість Цитоплазма впливає на реалізацію генетичної інформації хромосом, і тому є ше один аспект ролі цитоплазми в явищі спадковості — це так званий материнський ефект. Суть його полягає в тому, що властивості цитоплазми яйцеклітини формуються під контролем материнського генотипу. Перебудова (предетермІнація) властивостей цитоплазми яйця під впливом ядерних генів матері призводить до того, що властивості зиготи, ранні етапи розвитку, а іноді і весь онтогенез у значній мірі визначаються генотипом матері, а не зиготи. Це явище називається материнським ефектом і спостерігається тоді, коли цитоплазма в зиготу вноситься переважно яйцеклітиною. У більшості видів за запліднення саме жіноча гамета є донором цитоплазми, і тому материнський ефект спостерігається дуже часто. За дробління зиготи фенотипові ознаки матері виявляються до тих пір, поки цитоплазма клітин, що утворюються, не змінить своїх властивостей під впливом геному батька.

Найважливіші проблеми позахромосомної спадковості пов’язані з: 1) мітохондріями і пластидами як носіями генетичної інформації за цитоплазматичної спадковості; 2) успадковуванням ознак, що контролюються одночасно генами ядра і цитоплазми; 3) ендосимбіонтами, інфекційними агентами та деякими іншими позахромосомними елементи як носіями генетичної інформації; 4) предетермінацією генами ядра властивостей власне цитоплазми і материнським ефектом у спадковості.

Цитоплазматична спадковість Існує т.з. «материнський» тип успадкування, тому що зігота отримує гени цитоплазми в основному від яйцеклітини. Якщо буде зроблено рециплокне схрещування: кінь (самиця) х віслюк (самець) мул але якщо: віслюк(самиця) х кінь (самець) лошак Результати схрещувань дуже різні, тому що більшість цитоплазми отримує нащадок від матері. В цитоплазмі є мітохондрії та пластиди (у рослин), крім того в цитоплазмі яйцеклітини є велика кількість іРНК, повязаної з білком. Це є цілком материнська РНК та білок.

Мітохондріальна спадковість Існує припущення, що мітохондрії та пластиди — це трансформовані в процесі еволюції ендосимбіонти (бактерії), що ввійшли у взаємодію з попередниками сучасних клітин (симбіогенегична гіпотеза). Вони містять ДНК, де розташовани нехромосомні гени, які мають великий вплив на розвиток істоти. Гени хлорофілу знаходяться в пластидах (хлоропластах) і тому якщо нормальні хлоропласти не потрапили у майбутню яйцеклітину, то рослина буде білою, якщо потраплять пластиди нормальні, та ті що не можуть хлорофіл - то рослина буде строкатою, якщо тільки нормальні - забарвлення рослини теж буде нормальним.
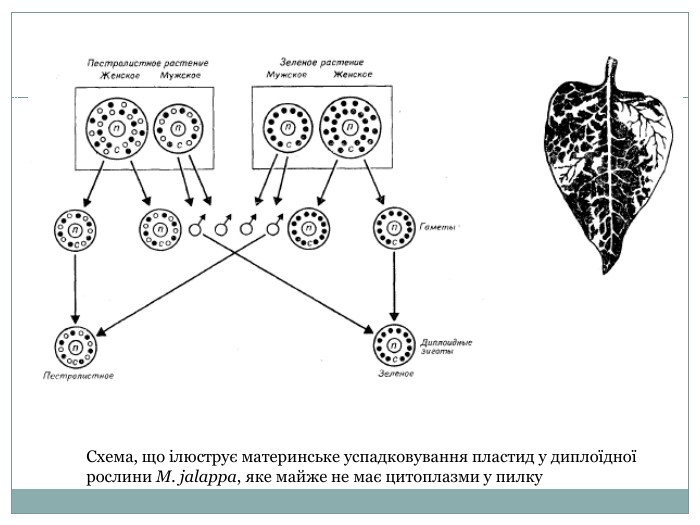
Схема, що ілюструє материнське успадковування пластид у диплоїдної рослини M. jalappa, яке майже не має цитоплазми у пилку
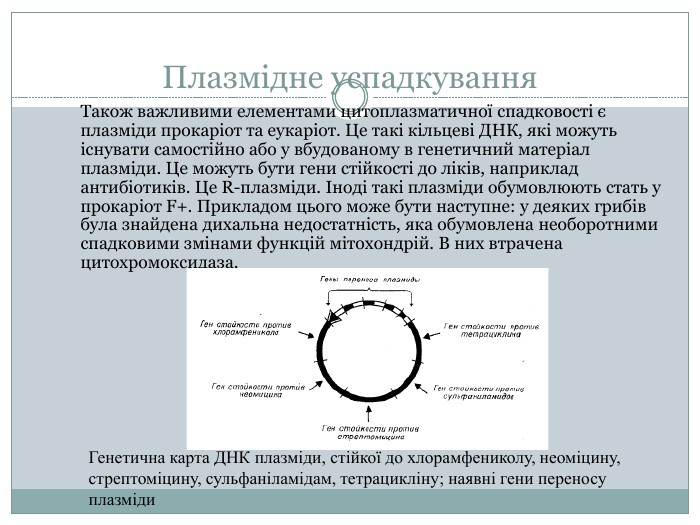
Плазмідне успадкування Також важливими елементами цитоплазматичної спадковості є плазміди прокаріот та еукаріот. Це такі кільцеві ДНК, які можуть існувати самостійно або у вбудованому в генетичний матеріал плазміди. Це можуть бути гени стійкості до ліків, наприклад антибіотиків. Це R-плазміди. Іноді такі плазміди обумовлюють стать у прокаріот F+. Прикладом цього може бути наступне: у деяких грибів була знайдена дихальна недостатність, яка обумовлена необоротними спадковими змінами функцій мітохондрій. В них втрачена цитохромоксидаза. Генетична карта ДНК плазміди, стійкої до хлорамфениколу, неоміцину, стрептоміцину, сульфаніламідам, тетрацикліну; наявні гени переносу плазміди

Були знайдені штами дріжджів, нейроспори мутанти по генам цитохромоксидази, які утворюють карликові колонії, в звязку з дихальною недостатністтю. Це вегетативні форми. Крім того були знайдена мутація при якій ген передається одним ядерним геном. Це карликовий штам. Тому при схрещувани можна спостерігати розщеплення аскоспор.

Цитоплазматична чоловіча стерильність Одних з найяскравіших прикладів успадкування чоловічої цитоплазматичної стерільності є успадкування у кукурудзи, цибулі, буряка, льону, томату та ін. У сучасному світі усі селекцеонери шукають гени чоловічої цитоплазматичної стерильності, тому що це дозволяє в багато разів скоротити термін виведення нових сортів рослин. Наприклад це відбувається з кукурудзою. Це дводомна культура, іноді чоловічі метелки знаходять порожнімі - спермії в них відсутні. Це успадковується по лінії рослин-матерів. У той же час жіночі рослини фертильні як завжди. Кукурудза має ознаку стерільності цитS. Відновлювач фертильності Rf- і відсутність генів rf-. Стерільною рослина може бути лише в тому випадку коли немає генів відновлювачів фертильності, але має ген цитS.
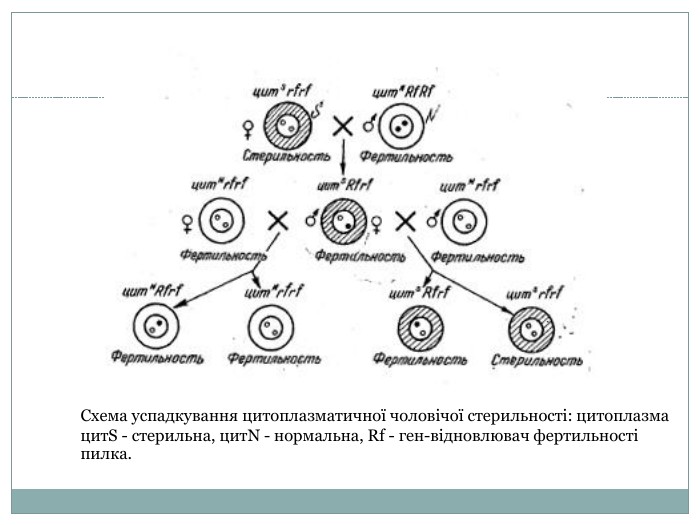
Схема успадкування цитоплазматичної чоловічої стерильності: цитоплазма цитS - стерильна, цитN - нормальна, Rf - ген-відновлювач фертильності пилка.
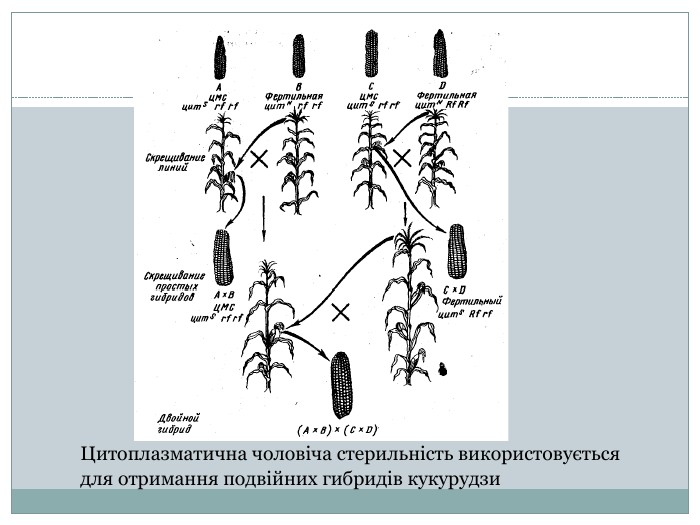
Цитоплазматична чоловіча стерильність використовується для отримання подвійних гибридів кукурудзи

Крітерії позахромосомного успадкування Відсутність менеделівського розщеплення на рівні тетрад, а також за аналізуючих та інших схрещувань Невідповідність результаів реципрокних зхрещувань Наявність материнського (іноді батьківського або змішаного) типу успадкування (фенотип нащадків визначається тим, кому із батьків належить цитоплазма зиготи) Незалежність успадкування ознак від наявності тих чи інших хромосом ядра; заміна в зіготі всіх батьківських хромосом материнськими чи навпаки не впливає на фенотип нащадків, в той час як заміна цитоплазми має вирішальне значення Наявність постзиготичних розщеплень, генетичної рекомбінації і вищеплення гаплоїдних сегрегантів за мітотичних поділів клітин, гетерозіготних по генах органел цитоплазми

У D. melanogaster відомі лінії без самців. Самки цих ліній при схрещуванні з будь-якими самцями дають у потомстві тільки самок. З'ясувалося, що лінії в яких выдсутні самці заражені спірохетами, які, проникаючи у відкладаються яйца, що, убивають чоловічі ембріони, але не вбивають жіночі ембріони. У результаті самки стають носіями інфекційного початку. Тісний зв'язок між функціями ядра й цитоплазматними ендосіимбіонтами був продемонстрований наприкінці 60-х років в Amoeba proieus К. Джеоном. Культура A. proteus була випадково заражена бактеріями, які проникнули в цитоплазму найпростішого й розмножувалися там до чисельності 150 тис. шт. на клітину. Більшість амеб загинулася, однак частина з них вижила й активно ділилася в присутності бактерії, що інфікувала їхню цитоплазму. Число бактеріальних клітин в амебі досягало тепер приблизно 50 000. Ядро найпростішого стало залежним від бактерії, що перетворилася в ендосимбионта. Пересаджене в іншу амебу ядро вже не могло забезпечувати функціонування й розподіл цитоплазми під час відсутності ніколи патогенних бактерій. Цей факт показує можливий шлях виникнення залежності генетичних функцій хазяїна від перебуває в його клітинах ендосимбионта. Факти такого роду використовують для підтвердження гіпотези про симбіогенетичного походження еукариотичної клітини, зокрема, про бактеріальне походження хлоропластів, мітохондрій і деяких інших клітинних органел, що містять ДНК, наприклад кинетосом у найпростіших. Уперше гіпотезу щодо походження хлоропластів висловив на початку століття професор Петербурзького університету А. С. Фамінцин (1835—1918). В останні роки в дрозофили виявлено кілька типів экстрахромосомных елементів, а також мігруючих елементів, які можуть міняти свою локалізацію в геномі. Вони одержали назву «стрибучих» генів. Ці елементи досить нагадують онкогенні Рнк-Утримуючі віруси, або ретровіруси. Ще більша подібність із онкогенними ретровірусами виявляють так звані мігруючі елементи генома дрозофилы, відкриті Д. Хогнессом, Г, П. Георгиевым, В. А. Гвозевым в 1976—1977 р. Їх називають множинними диспергованими генами (МДГ), оскільки вони повторюються в геномі розкидані по різних локусах. Їх називають також ретротранспозонами. Останнє найменування пов'язане з тим, що ці лементы можуть переміщатися по геному, і одним зі способів акой міграції є синтез на них РНК, а потім за допомогою зворотної траянскриптази синтез Днк-копії, впроваджуваної в нове місце. Для ряду МДГ показано, що в їхній склад, так само як до складу генома ретровирусов, входить ген, що кодує зворотну транскриптазу.

Предетермінація цитоплазми і материнський ефект Оскільки властивості цитоплазми яйця формуються під впливом ядерних генів матері, то фенотипи нащадків на ранніх стадіях їх розвитку, а Іноді і пізніше уподібнюються фенотипу матері (материнський ефект) Явише перебудови властивостей цитоплазми яйця (а отже і зиготи) під впливом ядерних генів матері називається генетичною предетермінацІєю цитоплазми. Якщо визвана зовнішніми умовами предетермінація цитоплазми призводить до досить тривалих змін фенотипу, які зберігаються іноді протягом кількох поколінь, то зазначені зміни називають тривалими модифікаціями.
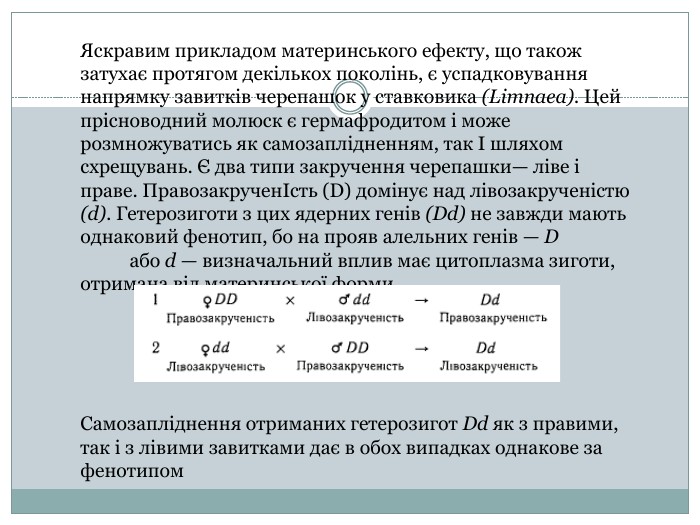
Яскравим прикладом материнського ефекту, що також затухає протягом декількох поколінь, є успадковування напрямку завитків черепашок у ставковика (Limnaea). Цей прісноводний молюск є гермафродитом і може розмножуватись як самозаплідненням, так І шляхом схрещувань. Є два типи закручення черепашки— ліве і праве. ПравозакрученІсть (D) домінує над лівозакрученістю (d). Гетерозиготи з цих ядерних генів (Dd) не завжди мають однаковий фенотип, бо на прояв алельних генів — D або d — визначальний вплив має цитоплазма зиготи, отримана від материнської форми Самозапліднення отриманих гетерозигот Dd як з правими, так і з лівими завитками дає в обох випадках однакове за фенотипом
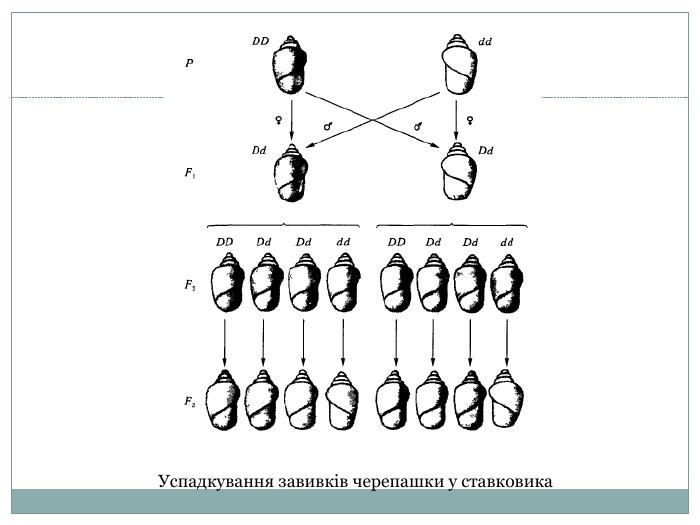
Успадкування завивків черепашки у ставковика
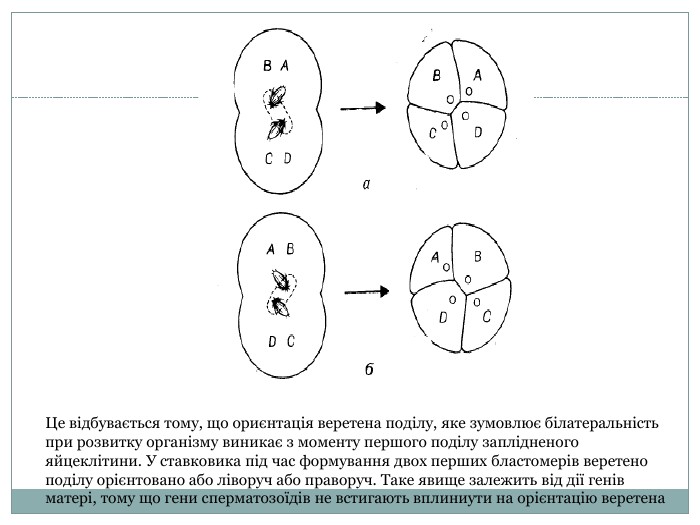
Це відбувається тому, що ориєнтація веретена поділу, яке зумовлює білатеральність при розвитку організму виникає з моменту першого поділу заплідненого яйцеклітини. У ставковика під час формування двох перших бластомерів веретено поділу орієнтовано або ліворуч або праворуч. Таке явище залежить від дії генів матері, тому що гени сперматозоїдів не встигають вплиниути на орієнтацію веретена

Вирішальну роль у цьому процесі грає генотип материнського організму, що визначає насамперед властивості цитоплазми яйцеклітини, від якого й залежить напрямок закручування раковини. Ця ознака формується в ранньому эмбриогенезі напрямком веретена другого дроблення. Таке явище одержало назву предетермінації цитоплазми генотипом матері або материнського ефекту.

Успадкування через інфекцію і ендосимбіонтів Існує лінія мишей з успадковуваною схильністтю до розвитку раку молочної залози, що передається по материнській лінії коли матір годує нащадків. Якщо до матері з ракових ліній які годують підсадти мишенят з неракової лінії, то такі мишата будуть схильні до раку молочної залози. Якщо мишенят ракової лінії вигодовують нормальні миши, то мишата будуть здорові. Таким чином ракова пухлина викликається впливом інфекційного фактору з молока матері (інфекційний фактор молока).

Успакдування через ендосимбіонтів У інфузорій є лінії які містять в цитоплазмі та виділяють у середовище специфічні частки, які мають назву каппа-частки. Носії каппа-часток є вбивці, і від цього не страждають, але парамеції інших ліній гинуть. Каппа-частики є бактерії (Caedobacter taeniospiralis), які є по відношенню до парамеції ендосимбіонтами. Вони містять білкову стрічку де знаходяться фаги - сімбіонти бактерій. Тут є тройний сімбіоз - інфузорія-бактерія-фаг. Якщо каппа-частка потрапляє в травну вакуоль інфузорії яка чутлива до цієї частки білкова стрічка бактерії розгортається, відповідно життєдіяльні фаги виробляють речовини, які спричиняють загибель інфузорій. Збереження каппа-часток в цитоплазмі та їх виведення з інфузорій-вбивців контролюється геном К. Рецесивна алель к не сприяє їх збереженню. Якщо відбувається прямий поділ то інфузорії-вбивці дають однотиповий клон із властивостями вбивць. Якщо відбувається схрещування в результаті конюгації двох клітин-вбивць можуть виникнути клітини без цих властивостей. Як результат автогамії ексконюгантів буде спостерігатись розщеплення за генотипом у співвідношенні 1 : 1 (КК : кк). Розподів каппа-часток буде залежити від тривалості конюгації батьківських клітин. Якщо конюгація дуже коротка каппа-частки не встигають потрапити в цитоплазма, якщо конюгація тривала, то ексконюгант від чутливої клітини отримує не тільки ген К і стає за генотипом Кк але і цитоплазму з каппа-частками. Таким чином все залежить чи є ген К і як довго продовжується конюгація.
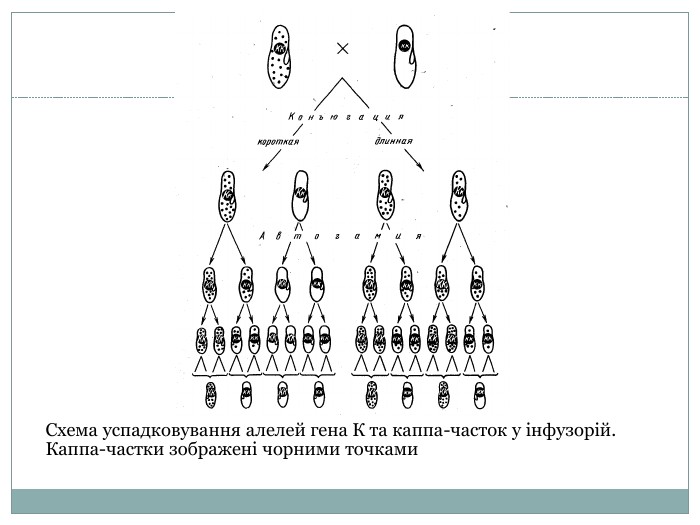
Схема успадковування алелей гена К та каппа-часток у інфузорій. Каппа-частки зображені чорними точками


про публікацію авторської розробки
Додати розробку