Амінокислоти їх будова та поширення .


Амінокислоти. Підготувала. Для учнів 10 класу Вчитель хімії Мандзюк Марія Іванівна

Мета: Розглянути основні поняття амінокислот, їх різновиди . Ознайомитися з хімічною будовою та ізомерією карбонового ланцюга, та оптичною ізомерією. Вивчимо функції та поширення амінокислот.
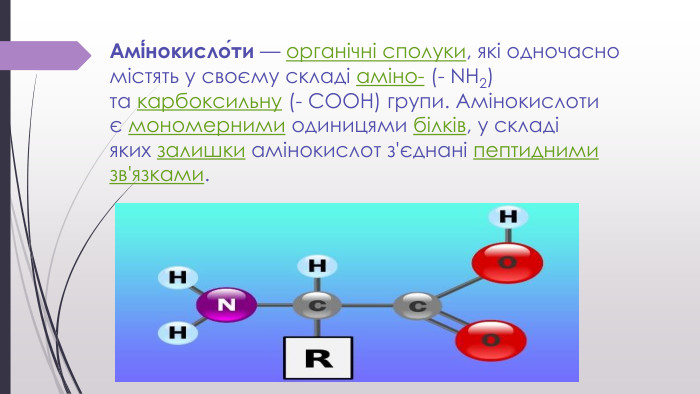
Амі́нокисло́ти — органічні сполуки, які одночасно містять у своєму складі аміно- (- NH2) та карбоксильну (- СООН) групи. Амінокислоти є мономерними одиницями білків, у складі яких залишки амінокислот з'єднані пептидними зв'язками.
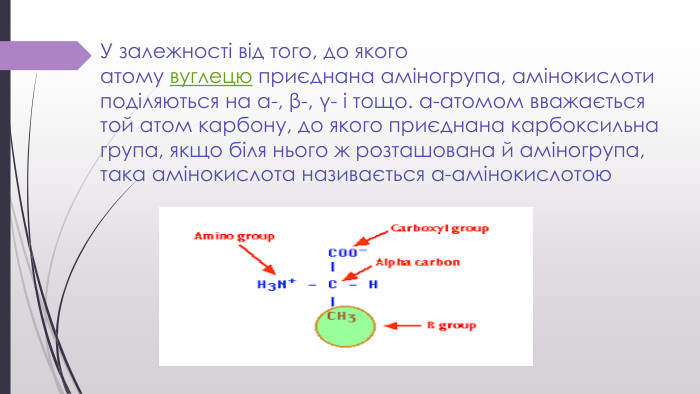
У залежності від того, до якого атому вуглецю приєднана аміногрупа, амінокислоти поділяються на α-, β-, γ- і тощо. α-атомом вважається той атом карбону, до якого приєднана карбоксильна група, якщо біля нього ж розташована й аміногрупа, така амінокислота називається α-амінокислотою

Номенклатура. Кожна з двадцяти стандартних, і багато нестандартних, амінокислот мають тривіальні назви. Частина цих назв пов'язана з джерелами, з яких уперше виділили сполуку: наприклад, аспарагін виділений зі спаржі (лат. Asparagus), глутамін — з глютену пшениці, тирозин — із сиру (сир грецькою tyros). Для скороченого запису протеїногенні амінокислоти позначають трибуквеним кодом, використовуючи перші три літери тривіальної назви (за винятком аспарагіну (Асн), глутаміну (Глн), ізолейцину (Іле) і триптофану (Трп), для останнього використовують також скорочення Три). Інколи також використовують позначення Asx і Glx, що означають «аспарагінова кислота або аспарагін» і «глутамінова кислота або глутамін». Існування таких позначень пояснюється тим, що під час гідролізу пептидів у лужних або кислих середовищах аспарагін і глутамін дуже легко перетворюються у відповідні кислоти, через що часто неможливо точно визначити, яка саме амінокислота була у складі пептиду без застосування особливих підходів.

Хімічна будова. Усі амінокислоти містять карбоксильну й аміногрупи. В α-амінокислот вони приєднані до одного і того ж атома карбону. Решту молекули називають бічним ланцюгом або R-групою. Ці групи відрізняються за розміром, формою, гідрофільністю, електричним зарядом, схильністю формувати водневі зв'язки і загальною реакційною здатністю надаючи кожній амінокислоті унікальних властивостей. У найменшої амінокислоти — гліцину — бічного ланцюга немає, біля α-атома карбону крім карбоксильної й аміногрупи розташовані два атоми гідрогену.


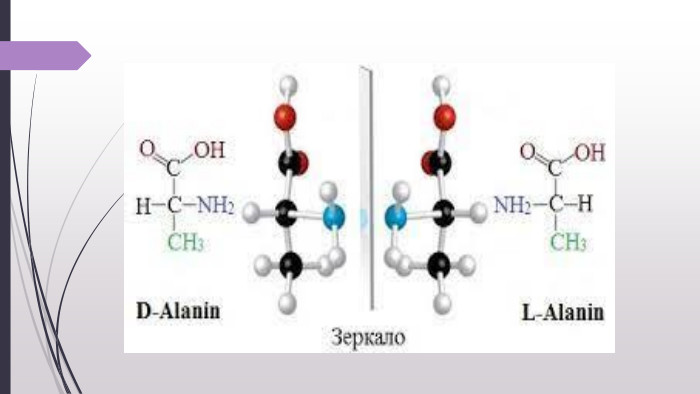
Оптична ізомерія. У всіх стандартних амінокислот, крім гліцину, α-атом карбону утворює ковалентні зв'язки із чотирма різними групами: карбоксильною, аміногрупою, бічним ланцюгом і атомом гідрогену. Отже, цей атом є хіральним центром. Таким чином, можливе утворення двох оптичних ізомерів, що є дзеркальними відображеннями одне одного. Вони не відрізняються за хімічними властивостями, але обертають площину поляризації світла в різні сторони. Такі ізомери називаються енантіомерами.

Для позначення абсолютної конфігурації чотирьох груп навколо хірального атома карбону використовується d/l система номенклатури розроблена Емілем Фішером. Він взяв за основу тріозу гліцеральдегід. Сполуки, які мають абсолютну конфігурацію, схожу до такої в l-гліцеральдегіду, називаються l-ізомерами, до d-гліцеральдегіду — d-ізомерами. Хоча історично літери l і d використовувались як скорочення від levorotatory (лівообертаючий) і dextrorotatory (правообертаючий), проте не всі l-амінокилсоти обертають площину поляризації світла вліво і навпаки.

Майже всі білки містять тільки l-ізомери амінокислот. d-амінокислоти знайдені в складі деяких поліпептидів, що утворюються в організмі морських черевоногих молюсків з роду Conus, та в складі клітинних стінок бактерій і деяких пептидних антибіотиків. Для формування стабільних повторюваних структур у білках необхідно, щоб усі амінокислоти, що входять до їхнього складу, були представлені тільки одним енантіомером — l або d. На відміну від звичайних хімічних реакцій, в яких переважно утворюються рацемічні суміші стереоізомерів, продуктами реакцій біосинтезу в клітинах є тільки одна із форм. Такого результату досягається завдяки ферментам, що мають несиметричні активні центри, а отже, є стереоспецифічними.

Класифікація на основі полярності бічних ланцюгівамінокислоти із неполярними бічними ланцюгами,амінокислоти із полярними незарядженими бічними ланцюгами (інколи розділяють на амінокислоти із неполярними аліфатичними та неполярними циклічними бічними ланцюгами),амінокислоти із позитивно зарядженими бічними ланцюгами,амінокислоти із негативно зарядженими бічними ланцюгами (інколи останні дві групи об'єднують в одну).

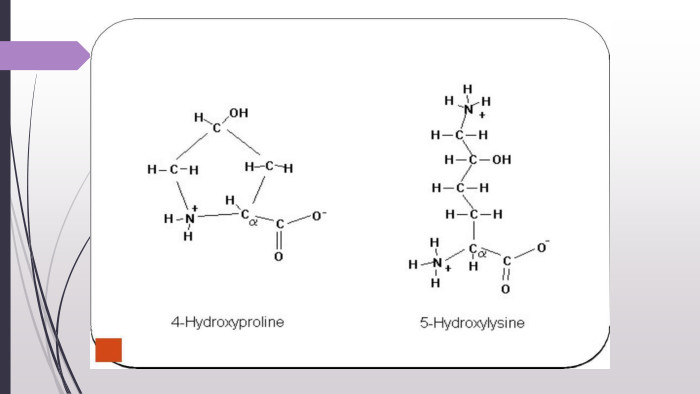
«Нестандартні» амінокислоти. Крім 20 «стандартних» амінокислот, що кодуються генетичним кодом[1], у живих організмах також трапляється велика кількість так званих нестандартних амінокислот. У складі готових поліпептидних ланцюгів деякі залишки амінокислот можуть модифікуватись. Таким чином, наприклад, утворюються нестандартні амінокислоти 5-гідроксилізин і 4-гідроксипролін. Обидві входять до складу білка сполучної тканини колагену, а гідроксипролін знайдений також у клітинних стінках рослин. 6-N-метиллізин є складовою скоротливого білка міозину. Деякі білки, які зв'язують іони кальцію, наприклад, протромбін, містять γ-карбоксиглутамат. Десмозин — складна амінокислота, що утворюється із чотирьох залишків лізину — знайдена у фібрилярному білку еластині. Багато залишків амінокислот можуть бути посттрансляційно модифіковані тимчасово з метою регуляції функції білків. До таких модифікацій належить приєднання фосфатних, метильних, ацетильних, аденільних, АДФ-рибозильних та інших груп.

Функції амінокислот. Деякі з 20 протеїногенних амінокислот називаються «незамінними» — це такі, що не виробляються в організмі і повинні надходити з їжею. Для людини це лізин, лейцин, ізолейцин, метіонін, фенілаланін, треонін, триптофан, валін, а для дітей також гістидин та аргінін.

Поширення амінокислот. Найпростіша амінокислота — гліцин — виявлена у міжзоряних хмарах та кометах. Амінокислоти виявлено і в метеоритах. Наприклад, у найкраще дослідженому метеориті Murchison виявлено понад 75 різних амінокислот, з них тільки 8 також знайдено в біологічних протеїнах. Ці амінокислоти мають виразно позаземне походження, багато з них не трапляються природно на Землі


Дякую за увагу!


про публікацію авторської розробки
Додати розробку