Презентація "Еволюція життєвих циклів водоростей, грибів та грибоподібних організмів".
Сформувати знання про походження водоростей, грибів та грибоподібних організмів; Ознайомити з особливостями існування водоростей, грибів та грибоподібних організмів.

 групи. Напряму підготовки Біологія Гуцол Наталія Миколаївна. Перевірила: доцент Журавель Н. М")
Міністерство освіти і науки України Національний Педагогічний Університет ім. Драгоманова Факультет Природничо-географічної освіти та екологіїКафедра біологіїТема: Еволюція життєвих циклів водоростей, грибів та грибоподібних організмів. Студентки 1 курсу, 1 Бм (З) групи. Напряму підготовки Біологія Гуцол Наталія Миколаївна. Перевірила: доцент Журавель Н. М

Водорості (лат. Algae) — гетерогенна група, деякі види автотрофні. Ця група охоплює кілька різних груп відносно простих за структурою живих організмів, які отримують необхідну для життєдіяльності енергію шляхом фотосинтезу, мешкають переважно у водному середовищі чи пристосувались до життя у ґрунті та інших наземних місцезростаннях. Водорості традиційно розглядалися як нижчі рослини і дійсно, деякі з них (Зелені водорості) близько споріднені до вищих рослин. Проте інші представляють різні групи найпростіших, наряду з тими, які традиційно вважалися схожими до тварин (наприклад, Apicomplexa). Різні групи водоростей виникли у різний час від різних предкових форм (поліфілетично) та розвивалися окремо, але в результаті конвергентної еволюції набули багато схожих рис.

Походження різних груп водоростей. Згідно з сучасною системою органічного світу, водорості є в обох надцарствах органічного світу: Procaryota та Eucaryota. Питання про походження водоростей тривалий час було дискусійним, власне, як і питання про походження еукаріот.
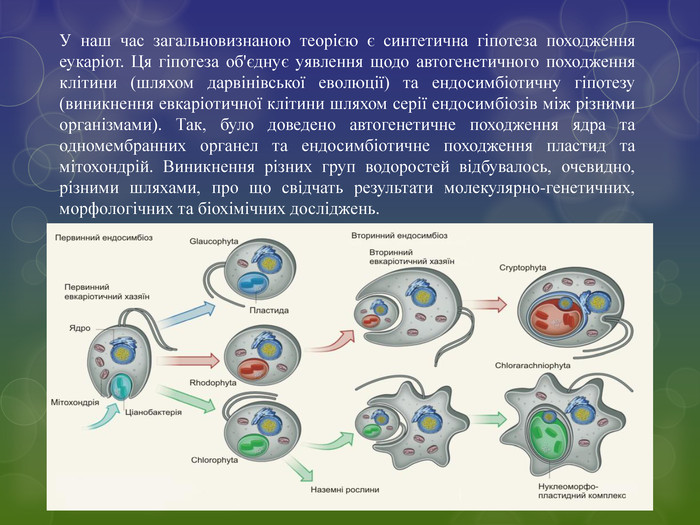
У наш час загальновизнаною теорією є синтетична гіпотеза походження еукаріот. Ця гіпотеза об'єднує уявлення щодо автогенетичного походження клітини (шляхом дарвінівської еволюції) та ендосимбіотичну гіпотезу (виникнення евкаріотичної клітини шляхом серії ендосимбіозів між різними організмами). Так, було доведено автогенетичне походження ядра та одномембранних органел та ендосимбіотичне походження пластид та мітохондрій. Виникнення різних груп водоростей відбувалось, очевидно, різними шляхами, про що свідчать результати молекулярно-генетичних, морфологічних та біохімічних досліджень.

Життєві цикли водоростей. Зміни життєвих форм, які відбуваються з індивідом протягом його онтогенезу, складають життєвий цикл. Життєвий цикл охоплює всі стадії розвитку індивіду між однаковими життєвими формами (наприклад, від зиготи до зиготи, або від гамети до гамети, від спорофіту до спорофіту). Розрізняють два основні типи життєвих циклів: життєві цикли водоростей, позбавлених статевого процесу, та життєві цикли водоростей, які мають статевий процес. Водорості, які позбавлені статевого процесу, називають агамними, а їх життєві цикли - цикломорфозом. У водоростей, що мають статевий процес - у так званих еугамних водоростей розрізняють три основні типи життєвого циклу - гаплофазний, диплофазний та гаплодиплофазний.
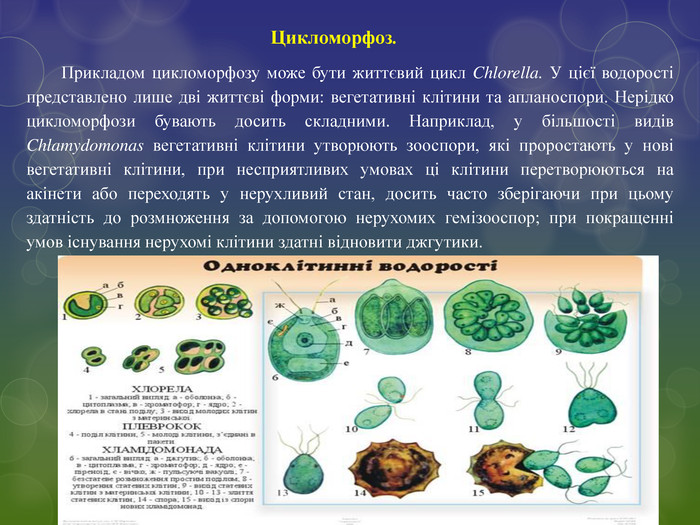
Цикломорфоз. Прикладом цикломорфозу може бути життєвий цикл Chlorella. У цієї водорості представлено лише дві життєві форми: вегетативні клітини та апланоспори. Нерідко цикломорфози бувають досить складними. Наприклад, у більшості видів Chlamydomonas вегетативні клітини утворюють зооспори, які проростають у нові вегетативні клітини, при несприятливих умовах ці клітини перетворюються на акінети або переходять у нерухливий стан, досить часто зберігаючи при цьому здатність до розмноження за допомогою нерухомих гемізооспор; при покращенні умов існування нерухомі клітини здатні відновити джгутики.
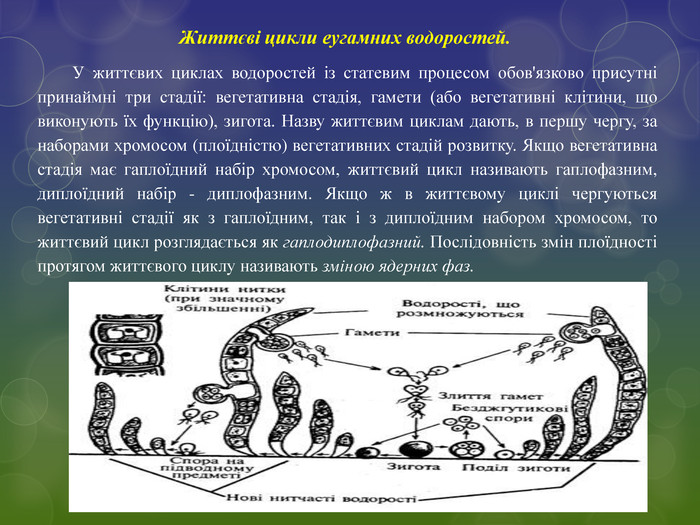
Життєві цикли еугамних водоростей. У життєвих циклах водоростей із статевим процесом обов'язково присутні принаймні три стадії: вегетативна стадія, гамети (або вегетативні клітини, що виконують їх функцію), зигота. Назву життєвим циклам дають, в першу чергу, за наборами хромосом (плоїдністю) вегетативних стадій розвитку. Якщо вегетативна стадія має гаплоїдний набір хромосом, життєвий цикл називають гаплофазним, диплоїдний набір - диплофазним. Якщо ж в життєвому циклі чергуються вегетативні стадії як з гаплоїдним, так і з диплоїдним набором хромосом, то життєвий цикл розглядається як гаплодиплофазний. Послідовність змін плоїдності протягом життєвого циклу називають зміною ядерних фаз.
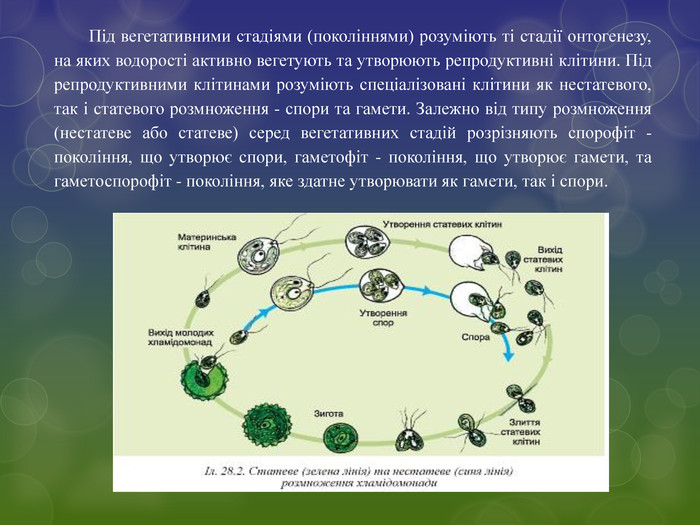
Під вегетативними стадіями (поколіннями) розуміють ті стадії онтогенезу, на яких водорості активно вегетують та утворюють репродуктивні клітини. Під репродуктивними клітинами розуміють спеціалізовані клітини як нестатевого, так і статевого розмноження - спори та гамети. Залежно від типу розмноження (нестатеве або статеве) серед вегетативних стадій розрізняють спорофіт - покоління, що утворює спори, гаметофіт - покоління, що утворює гамети, та гаметоспорофіт - покоління, яке здатне утворювати як гамети, так і спори.

У життєвому циклі може бути присутнє тільки одне покоління (наприклад, тільки гаметофіт або тільки гаметоспорофіт), або різні покоління нерегулярно чи регулярно змінюються одне одним (наприклад, спорофіт змінюється гаметофітом, а гаметофіт, у свою чергу - спорофітом). Залежно від цього розрізняють життєві цикли без зміни поколінь, з неправильним чергуванням поколінь та з правильним чергуванням поколінь. Якщо при чергуванні поколінь різні покоління морфологічно подібні (наприклад, гаметофіт зовні не відрізняється від спорофіту), то таке чергування поколінь називають ізоморфним, у протилежному випадку (наприклад, коли спорофіт макроскопічний, а гаметофіт мікроскопічний) чергування поколінь називають гетероморфним. Плоїдність поколінь, і, відповідно, тип життєвого циклу за чергуванням ядерних фаз, визначається тим, на якій стадії життєвого циклу відбувається мейотичний (редукційний) поділ. При мейозі кількість хромосом зменшується вдвічі. Таким чином, всі стадії розвитку протягом відрізку життєвого циклу від мейозу до статевого процесу будуть мати гаплоїдний набір хромосом, а всі стадії після статевого процесу до мейозу - диплоїдний набір.

Гриби́ (лат. Fungi) —царство еукаріотичних безхлорофільних гетеротрофних організмів, які живляться переважно осмотрофно, і більшість з яких здатні розмножуватись за допомогою спор (хоча деякі втратили цю можливість і розмножуються вегетативно). Більшість з них протягом всього життя або на певних стадіях розвитку мають міцеліальну будову, а деякі — дріжджі— одноклітинні. Сьогодні описано приблизно 70 тис. видів грибів, проте їх очікуване різноманіття, за оцінками різних авторів, становить від 300 тис. до 1,5 млн видів.
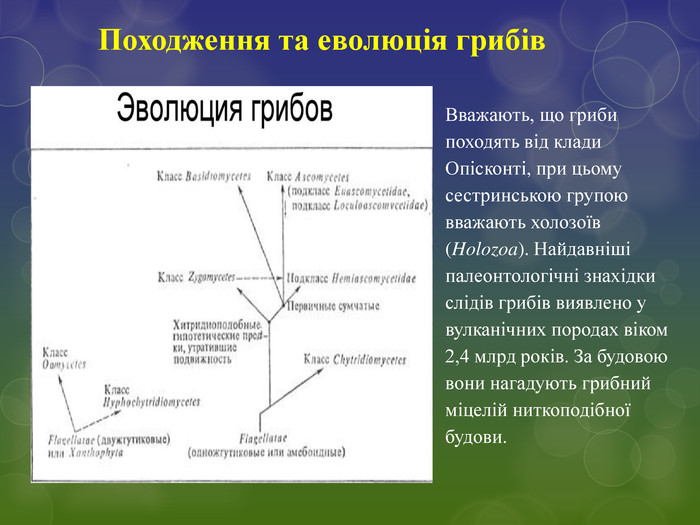
Вважають, що гриби походять від клади Опісконті, при цьому сестринською групою вважають холозоїв (Holozoa). Найдавніші палеонтологічні знахідки слідів грибів виявлено у вулканічних породах віком 2,4 млрд років. За будовою вони нагадують грибний міцелій ниткоподібної будови. Походження та еволюція грибів

Життєві циклиЄ єдиним типом життєвого циклу лише у акразіомікотових грибів. Здатність до статевого розмноження не виявлена або повністю втрачена у багатьох грибів, які за комплексом морфологічних, цитологічних та молекулярно-генетичних ознак належать до аскомікотових та базидіомікотових грибів. Оскільки системи цих відділів базують, в першу чергу, на ознаках, пов'язаних з органами статевого спороношення, для агамних представників не вдається визначити чіткого місця у системі цих відділів. Такі гриби об'єднані у штучну групу грибів з нез'ясованим систематичним положенням, яку сьогодні називають групою мітоспорових грибів, а також незавершеними грибами (Fungin Imperfecti) чи класом дейтероміцетів (Deuteromycetes). Мітоспорові гриби розмножуються виключно нестатевим шляхом, переважно за допомогою конідій. Таким чином, цикломорфоз є основним типом життєвого циклу у анаморфних грибів.

Хоча у більшості мітоспорових грибів статевий процес відсутній, рівень рекомбінантної мінливості є досить високим. Він обумовлений явищами гетерокаріозісу (різноядерності) та парасексуальним циклом. При гетерокаріозісі в різних клітинах одного міцелію можуть знаходитися генетично неоднакові ядра, які через анастомози переходять з однієї клітини міцелію в іншу. Хоча «мігруючі» ядра зазвичай не зливаються з ядрами клітини-реципієнта, але привносять нову генетичну інформацію і розширюють тим самим адаптивні можливості гриба по відношенню умов зовнішнього середовища.

В окремих випадках «мігруючі» ядра зливаються з ядрами клітини-реципієнта. Таке явище називають парасексуальним циклом. Воно призводить до утворення диплоїдних ядер, що забезпечують генетичні зміни грибного організму. Парасексуальний цикл включає кілька етапів: злиття ядер гетерокаріона і утворення диплоїдного гетерозиготного ядра; розмноження таких гетерозиготних ядер в міцелії; мітотична рекомбінація при розмноженні диплоїдних ядер; вегетативна гаплодізація диплоїдних ядер внаслідок втрати хромосом. Міцелій та спорангії різопусу чорніючого (Rhisopus nigricans) на поживному середовищі у чашці Петрі.
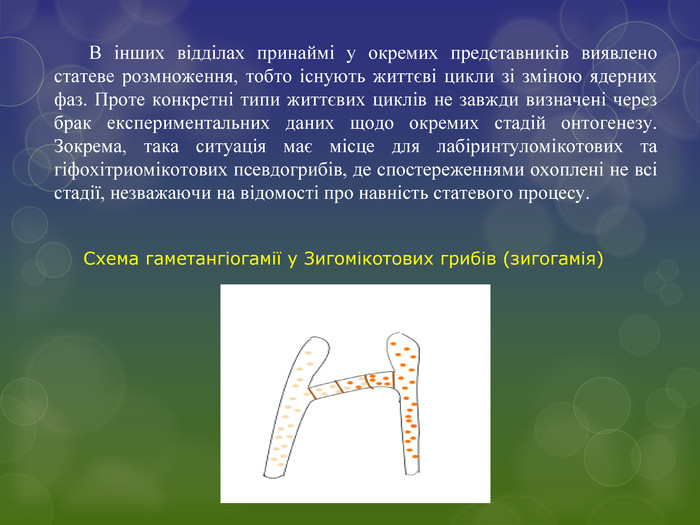
В інших відділах принаймі у окремих представників виявлено статеве розмноження, тобто існують життєві цикли зі зміною ядерних фаз. Проте конкретні типи життєвих циклів не завжди визначені через брак експериментальних даних щодо окремих стадій онтогенезу. Зокрема, така ситуація має місце для лабіринтуломікотових та гіфохітриомікотових псевдогрибів, де спостереженнями охоплені не всі стадії, незважаючи на відомості про навність статевого процесу. Схема гаметангіогамії у Зигомікотових грибів (зигогамія)

Життєві цикли евгамних грибів. В онтогенезі евгамних грибів наявна вегетативна стадії, представлена особинами з дикаріонтичними клітинами. Після статевого процесу утворюється не зигота, а клітина злиття з дикаріонтичними ядрами. Зигота у класичному розумінні цього терміну в онтогенезі може бути взагалі відсутня, хоча замість неї після каріогамії обов'язково утворються диплоїдні зиготичні ядра. Такі ядра є гомологами зиготи, і при розрахунку схеми життєвого циклу саме їх визначають як зиготу. Присутність у життєвому циклі грибів дикаріонтичних поколінь обов'язково відзначають у назві типу життєвого циклу. В межах конкретних відділів грибів життєві цикли за їх типом є переважно досить одноманітними, що дозволяє використовувати цю ознаку при характеристиці грибних таксонів найвищого рангу. Плазмодій Діктиостеліуму (Dictyostelum) під мікроскопом

Для міксомікотових характерні диплофазні життєві цикли з гаметичною редукцією та без зміни поколінь або гаплодиплофазні цикли зі споричною редукцією та неправильним чергуванням ізоморфних поколінь без дикаріонтичних фаз. У плазмодіофоромікотових життєві цикли гаплофазні, з зиготичною редукцією та чергуванням одноядерного гаметофіту та дикаріонтичного спорофіту. Оомікотові гриби мають диплофазні життєві цикли з гаметичною редукцією та без зміни поколінь. Колонія паразитичного гриба мадурели сірої (Madurella cinerea) на поживному середовищі у чашці Петрі. Цей гриб оселяється в тріщинах шкіри людини.

Для справжніх грибів у більшості випадків характерні життєві цикли з наявністю дикаріонтичних поколінь, і лише у частини хітридіомікотових та зигомікотових грибів такі покоління відсутні. Аскомікотові та базидіомікотові мають переважно гаплофазні життєві цикли з зиготичною редукцією та чергуванням міцеліїв з одноядерними та дикаріонтичними клітинами. У частини хітридіомікотових грибів життєвий цикл гаплофазний з зиготичною редукцією та чергуванням одноядерного плазмодію з літніми цистами та дикаріонтичного плазмодію з зимовими цистами. У іншої частини представників цього відділу життєвий цикл диплофазний з гаметичною редукцією та неправильним чергуванням поколінь і без стадій дикаріонів. Для зигомікотових характерні гаплофазні життєві цикли з зиготичною редукцією та без зміни поколінь.

Часто один і той же гриб в життєвому циклі може мати декілька типів спороношення: нестатеве та статеве, які послідовно змінюють одне одного. Таке послідовне чергування анаморф та тепломорф у грибів отримало назву плеоморфізму. Явище плеоморфізму широко представлено у грибів різних відділів, проте найбільшого розвитку воно отримало у аскомікотових. Строми аскомікотового гриба Elaphocordyceps ophioglossoides, який паразитує на оленячих трюфелях (Elaphomyces)
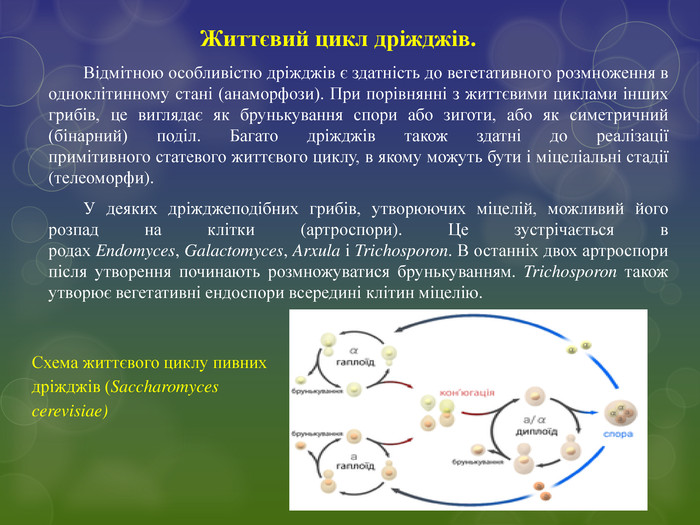
Життєвий цикл дріжджів. Відмітною особливістю дріжджів є здатність до вегетативного розмноження в одноклітинному стані (анаморфози). При порівнянні з життєвими циклами інших грибів, це виглядає як брунькування спори або зиготи, або як симетричний (бінарний) поділ. Багато дріжджів також здатні до реалізації примітивного статевого життєвого циклу, в якому можуть бути і міцеліальні стадії (телеоморфи). У деяких дріжджеподібних грибів, утворюючих міцелій, можливий його розпад на клітки (артроспори). Це зустрічається в родах Endomyces, Galactomyces, Arxula і Trichosporon. В останніх двох артроспори після утворення починають розмножуватися брунькуванням. Trichosporon також утворює вегетативні ендоспори всередині клітин міцелію. Схема життєвого циклу пивних дріжджів (Saccharomyces cerevisiae)

ДЯКУЮ ЗА УВАГУ.


про публікацію авторської розробки
Додати розробку