Матеріали до уроків біології 9 класу: посібник для розв'язання задач з генетики
Теоретичні та практичні матеріали , розв'язки задач , вправи для вчителів , учнів з генетики та молекулярної біології


Упорядник : Л. П. Дендюк , вчитель біології Глібівської ЗОШ І-ІІІ ступенів
Рецензент : Н. І. Савлюк , методист відділу освіти , молоді та спорту
Новоушицької територіальної громади
Методичний супровід щодо компетентнісного та діяльнісного підходу у розв’язанні задач з генетики , молекулярної біології та екології – Н. Ушиця , 2017 . – 67 с.
У посібнику міститься теоретичний матеріал , розв’язки задач, а також вправи для самостіцного розв’язування .
Призначений для вчителів біології ,екології , спеціалістів з охорони природи , учнів загальноосвітніх шкіл , абітурієнтів.
Зміст
Вступ ………………………………………………………………………….………….5
Алгоритм розв’язування задач…………………………………………….……………7
Моногібридне схрещування…………………………………………….………………8
Розв'язування задач на моногібридне схрещування при повному
домінуванні ознак………………………………………………………………………..9
Задачі для самостійного розв’язання..…….…………………………….……………..15 Дигібридне схрещування……………………………………………….………………16
Задачі для самостійного розв’язання…………………………………………………..20 Полігібридне схрещування………………………………….………………………….24
Задачі для самостійного розв’язання…………………………………………………..26
Приклади розв’язання задач на генетику статті……………………………………….28
Приклади розв’язування задач групи крові……………………………………………29
Контрльні завдання……………………………………………………………………...31
Синтез білка.Генетичний код…………………………………………………………...32
ДНК……………………………………………………………………………………….33
РНК……………………………………………………………………………………….40
Трансляція і транскрипція………………………………………………………………42
Регуляція біосинтезу білка………………………………………………………………48
Приклади розв’язання задач з молекулярної біології…………………………………50
Контрольні завдання ……………………………………………………………………57
Задачі з екології ………………………………………………………………………...58
Приклади розв’язання задач зекології………………………………………………….58
Додатки…………………………………………………………………………………...62
Література………………………………………………………………………………...67
Вступ
![]() – наука про живі організми, виникнення і розвиток
– наука про живі організми, виникнення і розвиток
живої природи, про загальні закони цього розвитку. Термін "біологія" у 1802 р. запропонували незалежно один від одного два вчені - французький Ж. Ламарк і німецький Г. Тревіранус.
Основними завданнями біології є пізнання процесів виникнення життя, вивчення історії розвитку живих організмів, їх онтогенезу та філогенезу. До біології належить велика кількість природничих наук, кожна з яких має свої завдання та властиві їй методи дослідження. Всі біологічні науки можна розподілити на дві основні групи: ботанічні, об'єктами дослідження яких є рослинні організми, та зоологічні, що досліджують тварин. Серед них у свою чергу можна виділити: систематику, морфологію, анатомію, гістологію, цитологію, фізіологію, генетику, селекцію, палеоботаніку і палеозоологію. В останній час виникли й успішно розвиваються нові розділи біології - біофізика, біохімія, радіобіологія, молекулярна біологія, космічна біологія та ін.
Останнім часом у загальній біології на основі великого фактичного матеріалу та результатів найновіших досліджень рослинних організмів відокремилась і успішно розвивається репродуктивна біологія, яка сформувалася на стику багатьох наук, зокрема ембріології, фізіології, карпології, цитології, генетики, селекції та ін.
Не претендуючи на повноту визначення поняття "репродуктивна біологія", наводимо наш підхід щодо розкриття цього терміну. Під репродуктивною біологією розуміємо сукупність генетичних, структурно-морфологічних і функціональних процесів у зв'язку з еколого - фізіологічними та віковими змінами, що обумовлюють перехід апікальних меристем від вегетативного до генеративного стану та утворення спеціалізованих статевих органів розмноження (М. Барна, 1989, 1997).
Зародженню репродуктивної біології сприяли такі відкриття другої половини XIX та першої половини XX століття, як відкриття законів спадковості та мінливості (Г. Мендель, 1865), подвійного запліднення у покритонасінних рослин (С. Навашин, 1898), закону гомологічних рядів у спадковій мінливості та вчення про центри походження культурних рослин (М. Вавілов, 1935). Саме вони спонукали вчених до поглибленого дослідження біології цвітіння, запилення і запліднення, розвитку зародка і насінини, спокою та проростання насіння, процесів успадкування при схрещуванні різних організмів, екологічних аспектів репродукції рослин, до перегляду існуючих і створення нових класифікацій та розробки нових концепцій щодо походження рослинного і тваринного світу зокрема та органічного світу загалом.
Це в свою чергу призвело до необхідності розробки нових методів та експериментальних підходів до вивчення морфолого-біологічних аспектів репродуктивного процесу живих організмів, глибшого пізнання їх генетичної природи. В цьому плані доцільно особливо виділити генетику, яка оформилась у самостійну науку.
Генетика — наука про спадковість і мінливість організмів; вивчає принципи зберігання, передачі й реалізації генетичної інформації. Як наука зародилась у 1900 році після перевідкриття голландським вченим Г. де Фрізом, німецьким вченим К. Корренсом і австрійським вченим Е. Чермаком законів успадкування ознак, відкритих у 1865 році чеським природодослідником Г. Менделем, і усвідомлення основоположного значення для біології. Вже в перше десятиріччя багато дослідників довели справедливість законів Мен-деля для найрізноманітніших організмів, що розмножуються статевим шляхом, і стала очевидною універсальність цих законів.
Термін "генетика" запровадив англійський вчений В. Бетсон (1906), а одиницю спадковості - ген (за Менделем, "елемент спадковості") - датський вчений В. Йогансен (1909).
Спадковість - властивість організмів повторювати в ряді поколінь подібні ознаки і забезпечувати специфічний характер індивідуального розвитку в певних умовах середовища.
Завдяки спадковості батьки і нащадки мають подібний тип біосинтезу, який визначає подібність у хімічному складі тканин, характері обміну речовин, фізіологічних відправленнях, морфологічних ознаках та інших особливостях.
Внаслідок цього кожний вид організмів відтворює себе із покоління в покоління.
Спадковість проявляється через розмноження, індивідуальний розвиток.
Мінливість - це явище, яке дещо протилежне спадковості, і виражається в тому, що в будь-якому поколінні окремі особини чимось відрізняються і одна від одної, і від своїх батьків. Мінливість - відмінність між особинами в межах виду. Мінливість проявляється за рахунок спадкових властивостей організму, зовнішніх умов.
Генетика є теоретичною основою селекції рослин, тварин і мікроорганізмів. Розвиток генетики людини призвів до розуміння того, що поряд із захворюваннями, які викликають бактеріальні, вірусні та інші інфекції, існує значне число (близько 2500) спадкових захворювань.
Генетична гетерогенність людської популяції включає цілий ряд аномалій обміну речовин, порушень конституції і психічних захворювань, причиною яких є генні мутації і хромосомні аберації.
Рання діагностика деяких спадкових захворювань дозволяє вчасно втрутитись у хід хвороби і дієтологічними або медикаментозними впливами запобігти аномальному розвитку і загибелі хворого. Так можна запобігти трагічним наслідкам і нормалізувати розвиток новонароджених, хворих галактоземією, які не засвоюють молочний цукор, або хворих фенілкетону-рією, чутливих до ароматичних амінокислот, якщо виключити з їхнього раціону небажані сполуки. Рання діагностика спадкових захворювань до народження дитини або визначення гетерозиготних носіїв генних і хромосомних аномалій сприяє запобіганню небажаних наслідків через планування сім'ї. Велику роль при цьому відіграє медикогенетичне консультування населення. До тих пір виліковування спадкових захворювань було неможливим. Всі розроблені заходи лише усували симптоми захворювань. Розвиток техніки генної інженерії в недалекому майбутньому обіцяє виникнення нової області медицини - генотерапії, завдяки якій можна буде виправити або замінити аномальні частини генетичного матеріалу.
Господарська діяльність людини часто пов'язана з втручанням у природні процеси, внаслідок чого скорочуються площі лісів, змінюється водний баланс, з'являються забруднюючі домішки у водоймах, повітрі та ґрунті. Прогнозування і запобігання можливих небажаних наслідків такого втручання неможливі без знання як екології, так і генетики, і насамперед генетики популяцій, яка оперує великими чисельностями організмів, що обмінюються генами в природних умовах.
При цьому необхідно передбачати збереження оптимальних розмірів і умов існування популяцій рослин, тварин і мікроорганізмів. Збереження їх генофонду - це збереження неоціненного природного багатства генів, які надалі можуть бути використані людиною в селекційному процесі. Дуже важливий аспект селекційної генетики - вивчення мутагенної активності різноманітних фізичних і хімічних агентів, що їх використовує людина. Поширення в нашому побуті мутагенів може підвищити концентрацію аномальних генів, збільшити вірогідність спадкових захворювань. Тому кожен новий вплив, кожна нова речовина, призначена для медицини, сільського господарства або харчової промисловості, проходять випробування на генетичну активність. Для цього генетики створюють спеціальні тест-системи: штами мікроорганізмів, культури дрозофіли, лінії мишей, культури клітин тварин і людини. І тільки переконавшись, що та чи інша речовина не мутаген,
1. Р – батьківські організми , узяті для схрещування ;
2. х - схрещування організмів ;
3. ♀ - жіноча стать ;
4. ♂- чоловіча стать ;
5. F - гібридні покоління (F1 - перше покоління ; F2 – друге покоління )
6. - особа чоловічої статті ;
7.
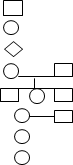 - особа жіночої
статті ;
- особа жіночої
статті ;
8. - стать невідома ;
9.
- шлюб та діти ( сибси );
10. - шлюб ;
11. --- -- - неофіційний шлюб ;
12. -==== - близькоспоріднений шлюб ;
можна використати її для тих чи інших цілей. Особлива важливість такої служби генетичної безпеки стає очевидною, якщо врахувати, що майже 90% мутагенів є канцерогенами Без перебільшення можна сказати, що генетика як наука про спадковість і мінливість знаходить застосування у всіх ділянках діяльності людини, пов'язаних з живими істотами: рослинами, тваринами і мікроорганізмами.

18.
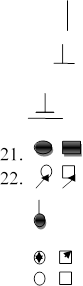 . - аборт ;
. - аборт ;
19. - медичний аборт;
20. - бездітний шлюб ;
- хворий ; - пробанд;
23. - мертвонароджений ;
24. - носій рецесивного гена ;
25. ! ! - особисто обмежений
Г. Мендель запропонував позначати ознаки літерами латинського алфавіту: А - ген домінантної ознаки організму; а - ген рецесивної ознаки організму.
Правила Г. Менделя, яких повинні дотримуватися всі генетики, використовуючи гібридологічний метод дослідження:
1.Схрещувані організми повинні належати до одного виду.
2.Схрещувані організми повинні чітко відрізнятися за окремими ознаками.
3.Досліджувані ознаки повинні бути константні, тобто відтворюватися з покоління в покоління при схрещуванні в межах лінії (батьківські форми).
4.Необхідні характеристика і кількісний облік усіх класів розщеплення, якщо воно спостерігається у гібридів першого і наступних поколінь.
Алгоритм розв’язування задач:
1. Прочитавши умову задачі, записати вихідні дані за допомогою загальноприйнятих символів.
2. Якщо перше покоління одноманітне, то фенотипічна ознака домінантна.
3. Якщо батьківські особини гомозиготи, то перше покоління одноманітне.
4. Чисті лінії (гомозиготи) дають завжди один сорт гамет.
5. Якщо особина має рецесивний фенотип, то вона рецесивна гомозигота (AA).
6. Гетерозиготи дають завжди парне число гамет, що визначається ступенем гетерозиготності даної особини (наприклад, тригетерозигота буде утворювати вісім типів гамет: 2 необхідно піднести до третього ступеня).
7. При схрещуванні гібридів завжди наявне розщеплення за досліджуваними ознаками, і, навпаки, якщо в поколінні є розщеплення, то батьківська (і) особина з домінантним фенотипом — гетерозигота.
8. При аналізуючих схрещуваннях число утворених у поколінні фенотипічних класів указує на число сортів гамет, утворених гібридом, причому всі фенотипічні класи будуть представлені рівними пропорціями
1 : 1 (1 : 1 : 1 : 1 і т. д.).
Моногібридне схрещування
Моногібридне схрещування - схрещування форм, при якому простежується успадкування за однією парою алелей, при цьому форми відрізняються однією протилежною ознакою. Г. Мендель проводив досліди на горосі, вибравши відмінні ознаки (вони не залежать від середовища і є альтернативними); простежував появу цих ознак у поколіннях; для схрещування використав особини, у яких у наступних поколіннях не було розщеплення ознак (чисті лінії); застосував математичний аналіз для обробки результатів; основні висновки одержані в дослідах з моногібридного схрещування.

Якщо материнська рослина мала червоні квітки, а батьківська — білі, то квітки всіх гібридних рослин виявляються червоними, біле забарвлення не проявляється. Значить, у гібрида Г1 з пари батьківських альтернативних ознак розвивається тільки одна, інша ознака не проявляється. Явище переважання у гібрида першого покоління ознак одного з батьків Мендель назвав домінуванням. Ознака, яка проявляється у гібрида і подавлює розвиток іншої альтернативної ознаки, була названа домінантною, подавлена — рецесивною. Це явище виявилось універсальним для рослин, тварин і людини й тому було виведено в ранг правила домінування. Оскільки всі гібриди першого покоління однакові, одноманітні, це явище стали називати першим законом. Менделя , або законом одноманітності гібридів першого покоління.
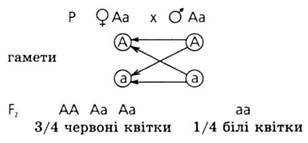
3 : 1 - формула розщеплення за фенотипом у F2 моногібридного схрещування при
повному домінуванні ознак.
1 : 2 : 1 - формула розщеплення за генотипом у F2 моногібридного схрещування при повному домінуванні ознак.
Якщо гібриду першого покоління надається можливість самозапилюватися, то в наступному поколінні, тобто в F2, з'являться рослини з ознаками обох батьків - з червоними і білими квітками.
Ця закономірність, яка полягає в появі у другому поколінні ознак обох батьківських організмів (домінантних і рецесивних), носить назву розщеплення. Розщеплення виявляється не випадковим, воно підкоряється певним кількісним закономірностям, а саме: 3/4 від загального числа рослин несуть червоні квітки і лише 1/4 - білі. Відношення числа рослин з домінантною ознакою до числа рослин з рецесивною ознакою виявляється рівним 3 : 1 . Значить, рецесивна ознака гібрида першого покоління не зникла, а лише була подавлена і проявилася в другому поколінні.
Розщеплення в F2 у певному кількісному співвідношенні домінантних і рецесивних ознак було названо законом розщеплення, або другим законом Менделя.
Другий закон Менделя: при схрещуванні двох гетерозиготних особин, тобто гібридів, які аналізуються за однією парою альтернативних видів ознаки, у потомстві спостерігається розщеплення за фенотипом у співвідношенні 3 : 1 і за генотипом 1 : 2 : 1.
Розв'язування задач на моногібридне схрещування при повному домінуванні ознак Приклади розв’язання задач
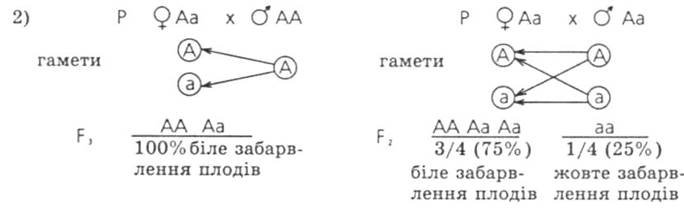
№1. Ген чорного забарвлення великої рогатої худоби домінує над геном чорного забарвлення. Яке потомство у F1 виникне від схрещування чорного гомозиготного бика з червоними коровами? Яким буде потомство від схрещувані F1 між собою таких гібридів? Які телята народяться від червоного бика і червоних корів з F?
Дано:
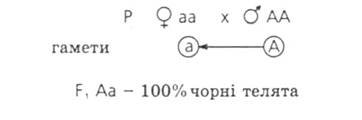 А — чорне забарвлення
А — чорне забарвлення
а — червоне забарвлення
 АА — чорне забарвлення Аа — чорне
забарвлення аа — червоне забарвлення
АА — чорне забарвлення Аа — чорне
забарвлення аа — червоне забарвлення
F1 -?

Відповідь: Потомство F1 - 100% чорного забарвлення. Серед потомства F 2 - 75% чорного забарвлення і 25% червоного забарвлення. Серед потомства F 3 - 50%
чорних і 50% червоних телят.
№2. У фігурного гарбуза біле забарвлення плодів домінує над жовтим. Якими будуть: 1) Гібриди F1 , F2, , якщо схрестити гомозиготний білий гарбуз з гомозиготним жовтим? 2) Нащадки від поворотного схрещування рослин F 1 з білою батьківською формою? 3) Нащадки від поворотного схрещування рослин F1 з жовтою батьківською формою?

№ 3. Голубоокий чоловік , обоє батьків якого мали карі очі , одружився з кароокою жінкою , в батька якої були карі очі , в матері –голубі . Від цього шлюбу
народився один голубоокий син. Визначіть генотип кожної цих осіб і складіть схему їх родоводу.
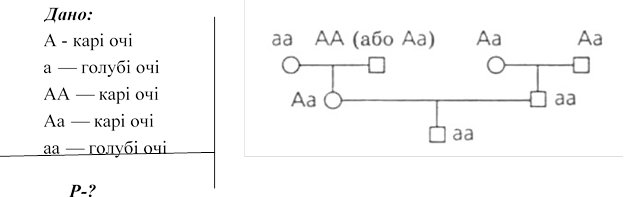
Відповідь: Чоловік гомозиготний рецесивний (аа), його обоє батьків гетерозиготні
(Аа), його дружина гетерозиготна (Аа), мати дружини гомозиготна рецесивна (аа), генотип батька дружини в цьому випадку встановити можна, він може бути гомозиготний домінантний (АА) або гетерозиготні (Аа), син цих людей гомозиготний рецесивний (аа).
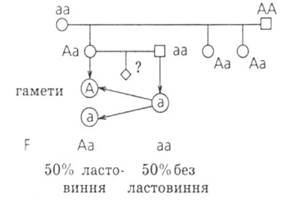
№ 4. Чоловік з ластовинням одружився з нормальною жінкою за цією ознакою. Від цього шлюбу народилося три дочки, які мають ластовиння. Одна з них одружилася з чоловіком без ластовиння. Які діти можуть народитися в цієї пари? Визначити, яка це ознака: домінантна чи рецесивна?
Дано:

 А – ластовиння а
— без ластовиння АА — ластовиння Аа — ластовиння аа — без ластовиння
А – ластовиння а
— без ластовиння АА — ластовиння Аа — ластовиння аа — без ластовиння
F -?
Відповідь: Наявність ластовиння - ознака домінантна. У цієї пар вірогідність народження дитини з ластовинням - 50%, без ластовиння - 50%
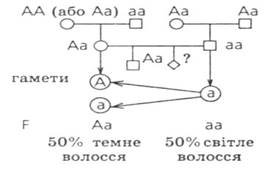
№ 5. Світловолосий юнак, батьки якого мали темне волосся, одружився темноволосою дівчиною, мати якої теж мала темне волосся, а батько - світле Від цього шлюбу у них народилася темноволоса дитина. Чи може народитися у них дитина зі світлим волоссям? Яка ймовірність народження такої дитині:
Дано:

 А - темне волосся а
— світле волосся АА — темне волосся Аа — темне волосся аа — світле волосся
А - темне волосся а
— світле волосся АА — темне волосся Аа — темне волосся аа — світле волосся
F -?
Відповідь: Вірогідність народження наступної дитини світловолосою в цій сім'ї - 50%.
№ 6 .Руде волосся - рецесивна ознака, неруде - домінантна. При яких умовах будуть народжуватись діти з імовірністю 100% з рудим волоссям? 50%? 25%?
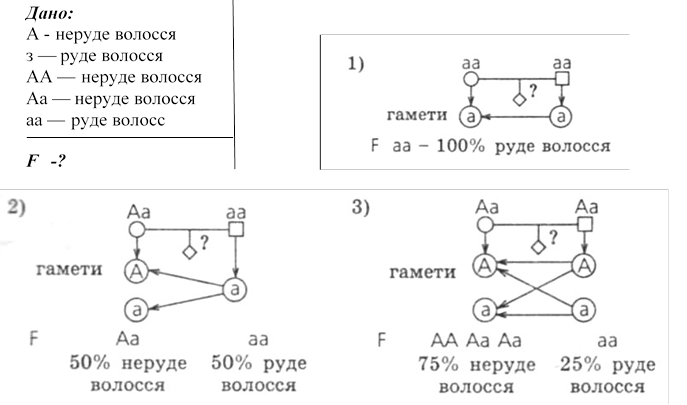
Відповідь: Будуть народжуватися діти тільки з рудим волоссям у сім'ї, обоє батьків гомозиготні рецесивні (аа) з рудим волоссям. З імовірністю - народження дитини з рудим волоссям можливе в сім'ї, де один з батьків гетерозиготний (Аа) за цією ознакою, а інший - гомозиготний рецесивний (аа). Ймовірність народження дитини з рудим волоссям становитиме 25% в сім'ї, де обоє батьків гетерозиготні (Аа) за цією ознакою.
№ 7. Альбінізм спадкується в людини як аутосомна рецесивна ознака. У сімї де один з подружжя альбінос, а інший - нормальний, народились різнояйцеві близнята, один з яких нормальний стосовно аналізованої хвороби, а інший альбінос. Яка вірогідність народження наступної дитини альбіносом?
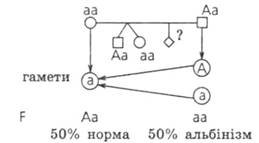 Дано:
Дано:
 А - норма з
— альбінізм АА — норма Аа — норма аа — альбінізм
А - норма з
— альбінізм АА — норма Аа — норма аа — альбінізм
F -?
Відповідь: Вірогідність народження наступної дитини альбіносом у цій сім'ї - 50%.
№ 8. У людини ген карого ока домінує над блакитним. Кароока жінка вийшла заміж за блакитноокого чоловіка. Який колір очей можливий у їхніх дітей?
 Дано: Розв'язок.
А - карий Р
♀ АА x ♂ аа а - блакитний
G А а
Дано: Розв'язок.
А - карий Р
♀ АА x ♂ аа а - блакитний
G А а
♀ АА F1 Аа - кароокі
♂ аа
F1 -?
Відповідь: 100% - одноманітне
№ 9. Гетерозиготна жінка вийшла заміж за гетерозиготного кароокого чоловіка.
Який колір очей можливий у їхніх дітей?
Дано: Розв'язок. А - карий Р ♀ Аа x ♂ Аа а - блакитний G А А
♀ Аа а а
 ♂
Аа F2
АА; Аа; Аа; аа
♂
Аа F2
АА; Аа; Аа; аа
F2 -?
Відповідь: за генотипом розщеплення 1:2:1, а за фенотипом 3:1.
№10. Який колір квітів буде у гібридів першого покоління білих та рожевих троянд по генотипу та по фенотипу, якщо білий колір - рецесивна ознака? (Схрещували гомозиготні особини).
 Дано: Розв’язок.
А – рожевий колір; Р
♀ аа х ♂ АА а – білий колір; G:
а А
Дано: Розв’язок.
А – рожевий колір; Р
♀ аа х ♂ АА а – білий колір; G:
а А
♂ АА F1 Аа
♀ аа
F 1 - ?
Відповідь: колір квітів буде рожевим.
№11. Якого кольору будуть троянди, одержані при схрещуванні двох гетерозиготних рослин рожевого кольору?
Розв’язок.
Р (F1)♀ Аа х ♂ Аа
G: А А а а
Відповідь: По закону розщеплення ознак, відбудеться розщеплення ознак: по генотипу у відношенні 1:2:1, по фенотипу 3:1. (аналізуюче схрещування).
Аналізуюче схрещування, його практичне значення.
№ 12. В курей нормальне оперення домінує над шовковистим. Є півень з нормальним оперенням невідомого походження. Як легше визначити його генотип?
Дано:
А – нормальне оперення Генотип може бути АА або Аа А – шовковисте оперення а) якщо:
АА - нормальне оперення Р→♂ ААнорм. х ♀ аашовк.
 аа - шовковисте оперення гамети → А → а генотипи ♂ норм. -
? F1→ Аа (100% нормальне оперення)
аа - шовковисте оперення гамети → А → а генотипи ♂ норм. -
? F1→ Аа (100% нормальне оперення)
б) Р →♂ Аа норм. х ♀ аашовк гамети → А а А а
F1→ Аа норм.; Аа норм.; аашовк.; аашовк. (50%:50% = 1:1)
Контрольні задачі.
Задача №1.
У вівса ген ранньостиглості домінує над геном пізньостиглості. Якого результату слід чекати від схрещування пізньостиглих рослин з гетерозиготними?
Задача №2.
У буряка від схрещування рослин з білими і червоними коренеплодами одержують рослини зі смугастими коренеплодами. Якого результату слід чекати від схрещування рослин, що мають смугасті і білі коренеплоди?
Задача №3.
У людини кароокістьвизначється домінантним геном, а голубоокість – рецесивним. Який колір очей можуть мати діти у кароокого батька і голубоокої матері?
Задача №4.
У людини ген полідактилії (шестипалість) домінує над нормальною будовою кисті. У сім’ї, де мати має нормальну кисть, а батько шестипалий, народилась дитина з нормальною будовою кисті. Які генотипи всіх трьох осіб?
Задача №5.
Дві чорні самки схрещувались з коричневим самцем. У однієї самки половина потомства була чорним, а половина коричневим, а в другої всі мишенята були лише чорними. Які генотипи самок, самця і потомства?
Задача №6.
Руде волосся – рецесивна ознака, неруде – домінантна. При яких шлюбах будуть народжуватись діти з імовірністю 100% з рудим волоссям? 50%? 25%? Складіть родоводи.
Задача №7.
Ген кароокості домінує над геном блакитноокості. Блакитноокий чоловік, батьки якого мали блакитні очі, одружився з кароокою жінкою, у батька якої були блакитні очі, а у матері - карі. Якого кольору очей можна очікувати у дітей цього шлюбу? Який генотип і фенотип вони матимуть? (р Аа х аа)
Задача № 8. Блакитноокий чоловік, обоє батьків якого мали карі очі, одружився з кароокою жінкою, у батька якої очі карі, а у матері - блакитні. Від цього шлюбу народився один блакитноокий син. Визначте генотипи кожного зі згаданих осіб.
Дигібридне схрещування - різновид схрещування, коли аналізується спадковість у потомстві двох ознак - гени ознак, які аналізуються, мають міститися в різних парах гомологічних хромосом
Приклади розв’язання задач
№1. Визначте генотип і фенотип кроликів від схрещування дигетерозиготної самки з дигомозиготним за рецесивними ознаками самцем, якщо чорний колір шерсті домінує над білим, а довгі вуха над короткими.
 Розв’язання:
Розв’язання:
Оскільки самка є дигетерозиготною, то її генотип — AaBb. Самець є дигомозиготним і має генотип — aabb.
Запишемо схему схрещування:
Р AaBb × aabb
♀ ♂
чорна білий
довговуха коротковухий
Жіноча особина може продукувати гамети типу AB, Ab, аB, аb, тоді як чоловіча — тільки гамети типу аb.
Позначимо типи гамет: G AB, Ab, аB, аb, аb.
Накреслимо решітку Пеннета і розв’яжемо задачу:
|
♂ ♀ |
ab |
ab |
ab |
ab |
|
AB |
AaBb чорні довговухі |
AaBb чорні довговухі |
AaBb чорні довговухі |
AaBb чорні довговухі |
|
Ab |
Aabb чорні коротковухі |
Aabb чорні коротковухі |
Aabb чорні коротковухі |
Aabb чорні коротковухі |
|
aB |
aaBb білі довговухі |
aaBb білі довговухі |
aaBb білі довговухі |
aaBb білі довговухі |
|
ab |
aabb білі коротковухі |
aabb білі коротковухі |
aabb білі коротковухі |
aabb білі коротковухі |
Отже, у потомстві одержано розщеплення 1 : 1 : 1 : 1 як за генотипом, так і за фенотипом.
Відповідь: у потомстві виявиться розщеплення ознак: за генотипами — AaBb, Aabb, aaBb, aabb у співвідношенні 1 : 1 : 1 : 1; за фенотипом — чорні довговухі, чорні коротковухі, білі довговухі й білі коротковухі у такому самому співвідношенні.
№ 2
У морських свинок ген чорного забарвлення домінує над геном коричневого забарвлення, а ген кошлатої шерсті домінує над геном гладенької. Який генотип і фенотип будуть мати нащадки від схрещування дигомозиготних особин?
Дано: Розв’язання:
 A — чорне
Оскільки і самка, й самець є дигомозиготними, то забарвлення генотип самки — AAВВ,
а генотип самця — aabb. a — коричневе
A — чорне
Оскільки і самка, й самець є дигомозиготними, то забарвлення генотип самки — AAВВ,
а генотип самця — aabb. a — коричневе
Запишемо схему схрещування:
забарвлення
Р ♀ AABB × ♂ aabb
В — кошлата шерсть
b — гладенька чорна коричневий шерсть кошлата гладенький
F1 — ?
Жіноча особина може продукувати тільки гамети типу AB, а чоловіча — тільки гамети типу ab.Позначимо типи гамет: G AB, ab.Накреслимо решітку Пеннета і розв’яжемо задачу
|
♂ ♀ |
ab |
|
AB |
AaBb чорні кошлаті |
Отже, всі нащадки будуть мати однаковий генотип AaBb, а за фенотипом усі вони будуть чорними з кошлатою шерстю.
Відповідь: усі нащадки будуть мати генотип AaBb і за фенотипом вони будуть чорними з кошлатою шерстю.
№ 3
У мухи-дрозофіли сіре забарвлення тіла домінує над чорним, а нормальна довжина крил домінує над короткою. Якими будуть нащадки від схрещування дигетерозиготних особин?
 Розв’язання:
Розв’язання:
Оскільки самка і самець є дигетерозиготними, то їхній генотип — AaBb.
Запишемо схему схрещування:
Р AaBb AaBb
♀
× ♂
сіра сірий
довгокрила довгокрилий
Жіноча особина, як і чоловіча, може продукувати гамети типу AB, Ab, aВ, ab.
Позначимо типи гамет: G AB, Ab, aВ, ab, AB, Ab, aВ, ab.
Накреслимо решітку Пеннета і розв’яжемо задачу:
|
♂ ♀ |
AB |
Ab |
aB |
ab |
|
AB |
AABВ сірі довгокрилі |
AABb сірі довгокрилі |
AaBВ сірі довгокрилі |
AaBb сірі довгокрилі |
|
Ab |
AABb сірі довгокрилі |
AAbb сірі короткокрилі |
AaВb сірі довгокрилі |
Aabb сірі короткокрилі |
|
aB |
AaBВ сірі |
AaBb сірі |
aaBВ чорні |
aaBb чорні |
|
|
довгокрилі |
довгокрилі |
довгокрилі |
довгокрилі |
|
ab |
AaВb сірі довгокрилі |
Aabb сірі короткокрилі |
aaВb чорні довгокрилі |
aabb чорні короткокрилі |
Отже, у потомстві одержано розщеплення за фенотипом — 9 : 3 : 3 : 1.
Відповідь: у потомстві відбудеться розщеплення ознак за генотипами AАBВ, AАBb, AaBВ, AaBb, AАbb, Aabb, aaBВ, aaBb, aabb. Кожна пара альтернативних варіантів ознак успадковується незалежно від інших пар і дає розщеплення 3 : 1 по кожній парі (як і при моногібридному схрещуванні). За фенотипами відбудеться розщеплення у співвідношенні 9 : 3 : 3 : 1, а саме: 9 сірі довгокрилі : 3 сірі короткокрилі : 3 чорні довгокрилі: 1 чорна коротко крила
№ 4.
Карі очі — домінантна ознака, блакитні — рецесивна. Темне волосся домінує над світлим. Темноволоса, дигетерозиготна жінка з карими очима одружилася із світловолосим чоловіком із блакитними очима. Визначте, який фенотип можуть мати діти в цій родині.
Орієнтир до розв’язання задачі:
1. Позначимо гени:
A — домінантний ген (темне волосся); а — рецесивний ген (світле волосся); B — домінантний ген (карі очі); b — рецесивний ген (блакитні очі).
2. Запишемо схему схрещування:
Р AaBb × aabb
♂
♀ ♂
темноволоса світловолосий кароока блакитноокий
Жіноча особина може продукувати гамети типу AB, Ab, аB, аb, а чоловіча — тільки аb.
Позначимо типи гамет: G AB, Ab, аB, аb, аb.
3.Накреслимо решітку Пеннета і розв’яжемо задачу:
|
♂ ♀ |
ab |
ab |
ab |
ab |
|
AB |
AaBb темноволосі кароокі |
AaBb темноволосі кароокі |
AaBb темноволосі кароокі |
AaBb темноволосі кароокі |
|
Ab |
Aabb темноволосі блакитноокі |
Aabb темноволосі блакитноокі |
Aabb темноволосі блакитноокі |
Aabb темноволосі блакитноокі |
|
aB |
aaBb світловолосі кароокі |
aaBb світловолосі кароокі |
aaBb світловолосі кароокі |
aaBb світловолосі кароокі |
|
ab |
aabb світловолосі блакитноокі |
aabb світловолосі блакитноокі |
aabb світловолосі блакитноокі |
aabb світловолосі блакитноокі |
Отже, у потомстві одержано розщеплення 1 : 1 : 1 : 1 як за генотипом, так і за фенотипом.
Відповідь: фенотип дітей становить: 50 % темноволосих, із них 25 % — кароокі й 25 % — блакитноокі та 50 % світловолосих, із них 25 % — кароокі й 25 % — блакитноокі. Контрольні задачі
Задача №1.
Гени чорної і короткої шерсті мишей домінують над генами білої і довгої шерсті. Схрестили гомозиготних чорних довгошерстих та білих короткошерстих мишей. Яким буде потомство в обох зворотних схрещуваннях?
Задача №2.
Які ознаки матимуть гібридні томати, одержані внаслідок запилення рослин із червоними матовими плодами, пилком жовтоплідних томатів із прозорою шкірочкою? Яке потомство отримаємо внаслідок схрещування таких гібридів? Червоний колір і матова шкірочка плодів – домінантні ознаки, а жовтий колір і прозора шкірочка плодів – рецесивні.
Задача №3.
У кролика звичайна шерсть домінує над подовженою ангорською, стоячі вуха – над капловухістю. Схрестивши кролика зі звичайною шерстю і стоячими вухами та ангорського капловухого, у потомстві одержали 25% кроликів зі звичайною шерстю і стоячими вухами, 25% - зі звичайною шерстю, але капловухих, 25% - ангорських зі стоячими вухами і 25% - ангорських капловухих. Які генотипи батьків і гібридів?
Задача №4.
У гороху гладенька форма насіння домінує над зморшкуватою, а високорослість – над низкорослістю. Обидві ознаки успадковуються незалежно. Схрестили дві гомозиготні рослини: високу зі зморшкуватим насінням і низькорослу – з гладеньким. Надалі гібриди розмножувалися самозапиленням. У F2 одержали 1760 рослин. Визначте серед них ймовірну кількість високих рослин із гладеньким насінням? Задача №5.
Чорна масть корів домінує над рудою, а білолобість – над суцільним забарвленням голови. Протягом кількох років схрещували рудих білолобих корів із чорним чорноголовим бугаєм. Одержали: чорних білолобих телят – 160, чорних чорноголових – 154, рудих білолобих – 158. Визначте ймовірну кількість рудих рудоголових телят. Задача №6.
У людини деякі форми короткозорості домінують над нормальним зором, а карі очі – над блакитними. Гени обох пар знаходяться в різних хромосомах.
1. Яке потомство можна чекати від шлюбу гетерозиготних за обома ознаками батьків?
2. Яке потомство можна чекати від шлюбу чоловіка з нормальним зором і блакитними очима і дигетерозиготної жінки ?
Задача №7.
Чоловік із кучерявим волоссям і карими очима, родичі якого в ряді поколінь мали карі очі, одружується з жінкою, яка має пряме волосся й блакитні очі. Чи будуть у них діти й онуки з кучерявим волоссям і блакитними очима за умови, що діти одружуватимуться з людьми, у яких блакитні очі й пряме волосся? Кучеряве волосся не сповна домінує над прямим, гетерозиготи мають хвилясте волосся. Задача №8.
Одружується дівчина, хвора на цукровий діабет і короткозорий юнак. Які у них можуть бути діти, якщо юнак гомозиготний? Чи будуть у них цілком здорові онуки, якщо їхні діти одружаться з особинами, хворими на цукровий діабет і нормальним зором? Цукровий діабет – рецесивна ознака, короткозорість – домінантна ознака.
Задача № 9
Ген раннього облисіння домінантний у чоловіка і рецесивний у жінки (ген знаходиться в аутосомі). Лисий гомозиготний чоловік без ластовиння одружився з жінкою, яка має густе волосся й ластовиння. Яка ймовірність народження дочок і синів без ластовиння і з раннім облисінням, якщо батько жінки був лисим гомозиготним і без ластовиння (ластовиння — домінантна ознака)?
Задача № 10
Серпоподібно-клітинна анемія успадковується як неповністю домінантна аутосомна ознака. Гомозиготні індивідууми вмирають зазвичай з настанням статевої зрілості; гетерозиготні— життєздатні, анемія у них виявляється субклінічно. Малярійний плазмодій не може використовувати для свого живлення с-гемоглобін. Тому люди, які мають цю форму гемоглобіну, не хворіють на малярію. Яка імовірність народження дітей, стійких до малярії, у сім'ї, де батько гетерозиготний щодо серпоподібно-клітинної анемії, а мати — здорова?
Задача №11
Таласемія успадковується як неповністю домінантна аутосомна ознака. У гомозигот це захворювання спричиняє смерть у 90-95% випадків, а в і гетерозит розвивається у відносно легкій формі. Яка вірогідність народження здорової дитини в сім'ї, де перша дитина й батько хворі на легку форму таласемії, а мати — здорова?
Задача №12
У медико-генетичну консультацію звернулось подружжя з питанням, чи можуть у них народитись здорові діти. У сім'ї було троє дітей. Двоє народилися мертвими з аненцефалією, а одна — з важкою клінічною картиною хвороби. Який прогноз можна дати щодо народження в цього подружжя здорової дитини?
Задача №13
Яка ймовірність народження здорової дитини в батьків, якщо аналізами у них і в їхньої першої дитини виявлено підвищений вміст цистину в сечі? Цистинурія — захворювання, що визначається рецесивним геном: у і гомозигот у нирках утворюються цистинові камені, а в гетерозигот спостерігається підвищений вміст цистину в сечі. Як успадковується цистенурія ? Задача № 14
Форма чашечки у квітці суниць може бути нормальною, зумовлені неповністю домінантним геном, і листкоподібною, що обумовлює його рецесивною алеллю. У гетерозигот чашечки мають проміжну форму. Визначте можливі генотипи й фенотипи рослин, отриманих схрещування гетерозигот з проміжною формою чашечки.
Задача №15
У людини серпоподібно-клітинна анемія — рецесивна ознака. Гомозиготи за рецесивним геном гинуть у ранньому віці від гемолітичної анемії. У гетерозигот захворювання розвивається в легкій формі. Визначте, яка ймовірність народження здорових дітей і дітей з легкою формою захворювання в батьків, які страждають легкою формою серпоподібно- клітинної анемії ?
Задача №16
Схрестили дві форми суниць: червоноплідну й білоплідну. Гібриди мають рожеві плоди. Якими за генотипом і фенотипом будуть рослини від схрещування між собою гібридів з рожевими плодами ?
Задача №17
Від схрещування двох форм нічної красуні одержали гібриди, 25% як мали червоні, 50% — рожеві, а 25% — білі квітки. Визначте генотипи і фенотипи батьківських рослин.
Задача №18
Батько глухонімий (рецесивна ознака) з гіпертонією (домінантна ознака), а мати здорова. У них народилась глухоніма дитина з гіпертонією. Чи можна стверджувати, що всі ці ознаки дитина успадкувала від батька ?
Задача № 19
Батько хворіє на мігрень (домінантна ознака), а мати здорова. У батька нормальний слух, у матері також, але вона має рецесивний алель глухоти. Яка ймовірність народження у них дитини з обома хворобами, якщо батько гетерозиготний за обома ознаками ?
Задача №20
Жінка з блакитними очима і темним волоссям вийшла заміж за кароокого чоловіка зі світлим волоссям. Відомо, що в батька дружини очі карі, а в матері — блакитні, батько дружини темноволосий, а мати — світловолоса. У батька чоловіка блакитні очі та світле волосся, у його матері — карі очі й темне волосся. Визначте генотипи згаданих осіб. Якими можуть бути діти від цього шлюбу?
Задача №21
У батька були карі очі й довгі вії, а в матері — сірі очі й короткі вії. Батько був гомозиготний за цими ознаками. Які ознаки будуть у їхніх дітей, якщо ці ознаки перебувають у різних хромосомах? Карі очі й довгі вії — ознаки домінантні. У яких шлюбах у їхніх онуків можуть народитися діти із сірими очима й довгими віями ?
Задача №22
Схрестили мишей, які мають чорне домінантне забарвлення шерсті й довгі вуха, з мишами, що мають рецесивне коричневе забарвлення шерсті та короткі вуха. Ген довгих вух є домінантним. Визначте відсоток генотипів серед потомства у F2, які відповідають генотипам батьків F1.
Задача № 23
У пшениці довге стебло і безосте колосся — домінантні ознаки, коротке стебло і остисте колосся — рецесивні. Схрестили пшеницю, яка має домінантні ознаки, із пшеницею, що має рецесивні ознаки. Який відсоток рослин з батьківськими генотипами утвориться в другому поколінні ?
Полігібридне схрещування
Аналіз схрещування одної пари ознак у моногібридному схрещуванні дозволяє зрозуміти спадкування двох і більше пар ознак при дигібридному і полігібридному схрещуваннях. Розщеплення в F2 за фенотипом для кожної пари альтернативних ознак рівне 3:1. Це вихідне відношення забезпечується точним цитологічним механізмом розходження гомологічних хромосом мейозі. Принцип незалежної поведінки різних пар альтернативних ознак у розщепленні за фенотипом у F2 виражається формулою (3+1)п, де n - ступінь гетерозиготності. Виходячи з наведеної формули, можна розраховувати число очікуваних класів у розщепленні за фенотипом при будь-якому числі пар ознак, взятих у схрещуванні: моногібридне схрещування (3 + І)1 =3:1, тобто 2 класи; дигібридне схрещування (3 + І)2 = 9 : 3 : 3 : 1, тобто 4 класи; тригібридне схрещування (3 + 1)3 =27:9:9:9:3:3:3:1, тобто 8 класів і т. д.
Інакше кажучи, число фенотипічних класів у F2 може бути виражено формулою 2n, де основа 2 вказує на парність (алельність) двох алелей одного гена, які знаходяться в одній парі гомозиготних хромосом, а ступінь n - число генів у негомологічних хромосомах, за якими відрізняються схрещувані батьківські форми. Таким же чином можна розрахувати число типів гамет, які утворюються в будьякого гібрида першого покоління, і число комбінацій гамет, даючи різні генотипи F2 , і в моногібрида Аа утворюються два сорти гамет (А і а), або 21; в дигібрида АаВв — чотири сорти гамет (АВ, Ав, аВ, ав), або 22; в тригібрида — вісім сортів гамет (АВС, АВс, АвС, Авс, аВС, аВс, авС, авс), або 23 і т. д.
Отже, число різних типів гамет, які утворюються гібридом F1 також може бути виражено формулою 2n, де n - число генів, за якими відрізняються схрещувані форми. Оскільки при моногібридному схрещуванні у гібрида F1, утворюються два сорти жіночих і чоловічих гамет, то очевидно, що при цьому можливе утворення чотирьох комбінацій у відношенні 1АА : 2Аа : 1аа, тобто 41. При дигібридному схрещуванні таких поєднань буде 42 = 16, при тригібридному - 43 = 64 і т. д., тобто число можливих комбінацій гамет виражається формулою 4 n, де основа 4 відображає число можливих комбінацій чоловічих і жіночих гамет у моногібридному схрещуванні, а n - число пар алелей.
Число генетичних класів у потомстві моногібрида складає 3, при дигібридному схрещуванні в F2 генотипних класів 9, або 32, при тригібридному - 33 і т. д. Отже, число генотипних класів можна визначити за формулою 3 n, де n - число гетерозиготних пар алелей.
Кількісні закономірності утворення гамет гібридами і розщеплення в їх потомстві при різних типах схрещування
|
Явище , що враховується |
Тип схрещування |
||
|
Моногібридне |
Дигібрид не |
Полігібридне |
|
|
Число типів гамет , що утворюються гібридом F1 |
2 |
22 |
2 n |
|
Число комбінацій гамет при утворенні F2 |
4 |
42 |
4 n |
|
Число фенотипів F2 |
2 |
22 |
2 n |
|
Число генотипів F2 |
3 |
32 |
3 n |
|
Розщеплення за фенотипом F2 |
3:1 |
(3:1)2 |
(3:1) n |
|
Розщеплення за генотипом F2 |
1:2:1 |
(1:2:1)2 |
(1:2:1) n |
Приклади розв’язання задач
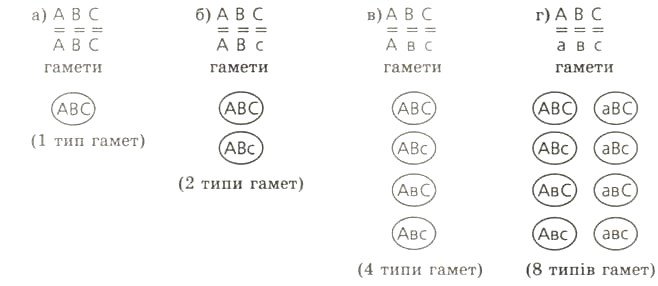
№1. Скільки і яких гамет може утворитися в самця морської свинки з чорним хутром, з довгою розеточною шерстю за умови: а) якщо він гомозиготний за всіма ознаками; б) якщо він гомозиготний за першими двома і гетерозиготний за третьою ознакою; в) якщо він гомозиготний за першою, але гетерозиготний за другою і третьою ознаками; г) якщо він гетерозиготний за всіма трьома ознаками. Поясніть це цитилогічно.
№2. У людини раннє облисіння домінує над відсутністю лисини у чоловіків, а у жінок воно рецесивне. Кароокий не лисий правша одружується з кароокою лівшою з густим волоссям. У них народилося троє дітей: кароокий син правша з раннім облисінням, блакитноока дочка правша з густим волоссям і кароокий син лівша з густим волоссям. Які генотипи батьків і дітей?
Дано :
 А- кароокість Аавв С1 С2
АаВв С2 С2 а-
голубоокість
А- кароокість Аавв С1 С2
АаВв С2 С2 а-
голубоокість
В- правша
В- лівша
 С1- раннє облисіння ААВв С1 С2 ааВв С1 С2 ААвв
С2 С2 С2-
відсутність (або АаВв С1 С2 ) (або
ааВв С1 С2 ) (абоАавв С2 С2)
раннього облисіння
С1- раннє облисіння ААВв С1 С2 ааВв С1 С2 ААвв
С2 С2 С2-
відсутність (або АаВв С1 С2 ) (або
ааВв С1 С2 ) (абоАавв С2 С2)
раннього облисіння
Відповідь: Генотип батька — АаВвС2С2, матері – АаввС1С2, кароокий син правша з раннім облисінням - ААВвС1С2, або АаВв С1С2блакитноока дочка правша з густим волоссям – ааВвС 1С2, або ааВвС2С2, кароокий син лівша з густим волоссям - ААввС2С2, або АаввС2С2.
Контрольні задачі
Задача № 1 . Які типи гамет утворюють особини з генотипом: а) AaBbCc, б) AaBBCc, в) AABbCC?
Задача № 2.
У рослини генотип AaBbCc. При самозапиленні цієї рослини (у випадку повного домінування за всіма парами алелів) скільки може бути в її потомстві: а) різних класів за фенотипом і їхнє співвідношення; б) різних класів за генотипом; в) можливих генотипів?
Задача №3.
Гетерозигота AABbCcDd схрещена з гомозиготним рецесивом aabbccdd. Визначте в потомстві від цього схрещування: а) число різних типів гамет; б) яка частина потомства буде мати всі чотири домінантні гени; в) яка частина потомства буде мати всі чотири рецесивні гени.
Задача №4.
У людини шестипалість (полідактилія), короткозорість і відсутність малих кутніх зубів передаються як домінантні аутосомні ознаки. Яка ймовірність народження дітей без аномалій у родині, де і батько, і мати мають усі три вади, але гетерозиготні за всіма трьома парами генів?
Задача №5 Короткозорий лівша одружується із жінкою, нормальною за обома ознаками. Відомо, що в подружжя були брати й сестри, які хворіли на
фенілкетонурію (нездатність засвоєння амінокислоти фенілаланіну), але самі вони нормальні щодо цієї ознаки. У їхніх родинах перша дитина була нормальною стосовно всіх трьох ознак, друга була короткозорою лівшею, а третя — хворою на фенілкетонурію. Визначте генотипи батьків і всіх трьох дітей, якщо відомо, що короткозорість — домінантна ознака, ліворукість і фенілкетонурія — рецесивні ознаки
Задача №6. Деякі форми катаракти і глухонімоти в людини передаються як аутосомні рецесивні не зчеплені між собою ознаки. Відсутність різців та ікол верхньої щелепи також може передаватись як рецесивна ознака, не зчеплена з катарактою і глухонімотою, а) Яка вірогідність народження дітей з усіма трьома аномаліями в сім’ї, де обоє батьків гетерозиготні за всіма трьома парами генів? б) Яка вірогідність народження дітей з всіма трьома аномаліями в сім’ї, де один з батьків страждає катарактою і глухонімотою, але гетерозиготний за третьою ознакою, а інший - гетерозиготний за катарактою і глухонімотою, але страждає відсутністю різців та ікол у верхній щелепі?
Задача №7. У горошку запашного висота Т домінує над висотою карликових рослин t, зелені боби C - над жовтими c, а опукле насіння Z - над зморшкуватим z. Який фенотип матимуть F1 і F2 від схрещування гомозиготного карликового зеленого горошку з гомозиготним високим жовтим опуклим горошком?
Задача №8 У курей ноги з пір’ям (Р) домінують над голими (p), трояндовидний гребінь (К) - над простим (k), а пір’я леггорнів (І ) - над забарвленим пір’ям (і). а) Курка з пір’ям на ногах, простим гребенем і білим пір’ям схрещена з півнем із пір’ям на ногах, трояндовидним гребнем і забарвленим пір’ям. Визначіть розщеплення в наступному поколінні за фенотипом. (Курка дигібридна, півень дигібридний.) б) Курка з пір’ям на ногах, трояндовидним гребенем і білим пір’ям схрещена з півнем, який має голі ноги, простий гребінь і забарвлене пір’я. Одне з курчат, одержаних від цього схрещування, мало всі ознаки півня. Чи можна встановити генотип курки?
Задача №9.У томатів червоне забарвлення плодів зумовлено домінантним геном У, а жовте - його рецесивнем алелем у, високі стебла — домінантним геном D, карликовість його рецесивним алелем d, гладенька шкірка плода — домінантним геном Н, опушена — його рецесивним алелем h. Схрещується рослина, гомозиготна за ознаками червоного забарвлення плодів, високого стебла і гладенької шкірки з карликовою рослиною, яка має жовті опуклі плоди, а) Визначіть генотип батьківської рослини, б) Визначіть генотип і фенотип потомства в F1. в) Яка частина потомства в F2 матиме жовті опушені плоди і карликовий зріст? г) Яка частина потомства в F2 матиме червоні гладенькі плоди і високе стебло? д) Яка частина потомства в F2 матиме червоні гладенькі плоди і карликовий зріст? е) Яка частина потомства в F2 буде гетерозиготна за всіма трьома парами алелей? ж) Яка частина потомства в F2 буде гомозиготна за всіма трьома парами алелей?
Приклади розв’язання задач на генетику статі
Задача 1.Які будуть кошенята від рудого кота та темної гетерозиготної кішки ?
Дано:

Розв’язок.
Р ♂ Хb Y x ♀ XBXb
|
|
Хb Y |
|
XB Xb |
XBХb темна кішка XBY темний кіт XbХb руда кішка Xb Y рудий кіт |
Задача 2. Чоловік, хворий на гемофілію, одружується зі здоровою жінкою, батько якої потерпав від гемофілії. Визначте ймовірність народження в цій родині здорових дітей.

Відповідь. 50 % того, що у цієї сімейної пари будуть діти страждати на гемофілію
Приклади розв’язання задач на групи крові
Позначення основних круп крові за системою АВО.
І група крові - ІОІО;
ІІ група крові - ІАІА; ІАІО; ІІ група крові - ІВІВ; ІВІО; ІV група крові - ІАІВ.
№. 1. В сім’ї, де в батька ІV група крові, а у матері – ІІ група, народилося 4 дітей з І, ІІ, ІІІ, ІV групами крові. Судмедекспертиза встановила, що один з дітей позашлюбний. Встановіть генотипи батьків, та визначте, яка група крові у позашлюбної дитини.
Запишемо коротку умову задачі:
♀ - ІАІО або ІАІА - ІІ група крові.
♂ - ІАІВ - ІV група крові.
F1: ІОІО- І група крові.
ІАІО або ІАІА - ІІ група крові.
ІВІО або ІВІО - ІІІ група крові.
ІАІВ - ІV група крові.
Визначимо, яка дитина у батьків не рідна. Перший випадок:
Р: ♀ ІАІО х ♂ ІАІВ G: ІА ; ІО ; ІА ;ІВ.
F1: ІАІА- ІІ група крові.
ІАІВ - ІV група крові.
ІАІО - ІІ група крові.
ІВІО - ІІІ група крові. Другий випадок: Р: ♀ ІАІА х ♂ ІАІВ G: ІА ; ІА ; ІА ;ІВ.
F1: ІАІА- ІІ група крові.
ІАІВ - ІV група крові.
Відповідь: у батьків з ІІ та ІV групами крові позашлюбна дитина з І групою крові
ІОІО.
№ 2. Відомо, що мати дівчинки з ІІІ групою крові (Rh-) мала І групу крові (Rh+). У двох її синів кров ІІ групи (Rh-). Встановити генотип і фенотип батька за трупою крові та резус-фактором.
Запишемо коротку умову задачі: Д – ген Rh+ d – ген Rh-
ДД або Дd – генотип Rh+ dd – генотип Rh-
F1 - ІВІО або ІВІВ – (ІІІ група крові, Rh-).
♀ (мати) – ІОІО – (І група крові, Rh+)
F1 (сини) – ІАІО або ІАІА – (ІІ група крові, Rh-).
29
Визначити генотип і фенотип батька -?
Розв’язання:
Так як в матері О група крові, а у її дітей групи А і В, то в батька дітей повинна бути ІV група крові ІАІВ.
Так як в матері Rh+, а в дітей Rh-, то в батька повинен бути Rh- Р: ♀ Дd ІОІО х ♂ dd ІАІВ
G: Д ІО ; d ІО d ІА ; d ІВ
№ 3 .У матері I група крові, у батька група крові III. Чи можуть діти успадкувати групу крові матері? Які генотипи матері та батька?
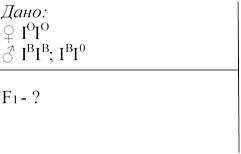 Розв’язок 1. Р ♀ ІОІО x♂ ІВІВ
Розв’язок 1. Р ♀ ІОІО x♂ ІВІВ
G ІО ІВ
F1 ІВ ІО
2. Р ♀ ІОІО x♂ ІВІО
G ІО ІВ ІО
F1 ІВ ІО; ІОІО
Відповідь: Діти можуть успадкувати групу крові матері лише в тому випадку, якщо генотип батька ІВІ0.
30
№ 4. У пологовому будинку переплутали двох хлопчиків. Батьки одного з них мали групи крові А і О, батьки іншого — А і АВ, хлопчики мають групи крові А і О.
Визначте, де чий син та генотипи батьків і дітей.
Скорочений запис умови задачі: Перша пара: Друга пара: мати АА(АО); мати АА(АО); батько ОО. батько АВ Хлопчики: АА(АО), ОО.
Розв’язок. Відомо, що генотип людей з групою крові О – іОіО, з групою крові АВ – генотип ІАІВ, з групою крові А – генотип ІАІО, або ІАІА.
Можливі генотипи дітей першої подружньої пари:
Р ♀ ІАІА × ♂ іОіО Р1 ♀ ІАіО × ♂ іОіО
F1 4 ІАіО – ІІ гр. 𝐹11 2 ІАіО – ІІ гр.: 2 іОІО – 1 гр.
Можливі генотипи дітей другої подружньої пари:
Р ♀ ІАІА × ♂ ІАІВ Р1 ♀ ІАіО × ♂ ІАІВ F1 ІАІА, ІАІВ, ІАІВ, ІАІА𝐹11 ІАІА, ІАІВ, ІАІО, ІВІО.
ІІ гр., IV гр., ІІ гр., IV гр. ІІ гр., IV гр., ІІ гр., ІІІ гр.
У другої подружньої пари не може бути дітей з першою групою крові, тобто хлопчик з групою крові О належить першій подружній парі, а хлопчик з групою крові А — другій.
Відповідь. Хлопчик з групою крові О належить першій подружній парі, а хлопчик з групою крові А — другій.
Контрольні завдання Задача 1.
Жінка гетерозиготна за А (ІІ) групою крові вийшла заміж за чоловіка з АВ (ІV) групою крові. Які групи крові матимуть їхні діти?
Задача 2.
За якого генотипу та фенотипу батьків діти не можуть успадкувати групу крові батька або матері?
Задача 3.
Від шлюбу між чоловіком з групою крові АВ і жінкою з групою крові А народилося троє дітей з групами крові В, АВ і О. Визначте генотипи батьків і дітей. Чи немає в цих даних чогось такого, що викликає сумнів?
Задача 4.
У подружжя було встановлено позитивний Rh – фактор, але в них народився хлопчик з негативним Rh – фактором. Як успадковується Rh – фактор?
Задача 5.
Під час розгляду судової справи про батьківство встановлено, що в дитини ІV група крові, у матері – ІІ група, а ймовірного батька – І група крові. До якого висновку повинен прийти судмедексперт? Задача 6.
Жінка, що має резус-позитивну кров ІІ групи, батько якої мав резус-негативну кров І групи, одружилась з чоловіком, який має резус-негативну кров І групи. Яка ймовірність народження в цій сім’ї дитини, яка успадкує обидві ознаки батька?
Задача 7.
Чоловік з резус-негативною І групою крові одружився з жінкою, що має резуспозитивну ІV групу крові. Які діти за групами крові та резус-фактором можуть у них народитись?
Задача 8.
Жінка з ластовинням (домінантна ознака) і ІІ групою крові вийшла заміж за чоловіка з І групою крові без ластовиння. Троє дітей від цього шлюбу не мали ластовиння, а одна дитина народилася з І групою крові. Визначте генотипи батьків і дітей.
Синтез білка. Транскрипція і трансляція. Генетичний код
1868-1869 роки - Йоган Фрідріх Мішер, працюючи в Тюбінгенській лабор; торії Е. Ф. Гоппе - Зайлера, досліджував клітини гною із лейкоцитів і сперматозоїди лосося і виділив речовину з сильними кислими властивостями, яку назва нуклеїном. Нуклеїн виявився тісно пов’язаним з основною білковоподібною речовиною, яку Мішер у 1872 році відділив від нуклеїну і назвав протаміном.
1889 рік - Р. Альтман вводить термін “ нуклеїнова кислота ”.
Починаючи з 1889 року, А. Коссель став проводити свої дослідження хімії нуклеїну. Він показав, що до його складу, крім фосфорної кислоти входять пурини, піримідини (азотисті основи, а також вуглеводневі компоненти). Було виявлено чотири азотисті основи: дві пуринові - аденін і гуанін, та дві піримідинові - тимін і цитозин.
У роботах 1900-1932 рр. А. Левене вияснив основні хімічні принципі будови вуглеводів нуклеїнових кислот, які відрізняються від глюкози, фруктози і галактози тим, що вони містять п’ять атомів вуглецю замість шести. Він виявив, що одна група вуглеводів нуклеїнових кислот містить на один атом кисню менше. А. Левене назвав групу з таким вуглеводом дезоксирибозою, а іншу - рибозою. На основі цього перші були названі дезоксирибонуклеїновими кислотами, а другі - рибонуклеїновими кислотами.
1924 рік - Р. Фельген розробив методи цитологічного розпізнавання ДНК і РНК. Він цитологічно довів, що ДНК локалізується в ядрах клітин, а РНК - у цитоплазмі.
1928 рік - Ф. Гріффіт відкрив у природі явище трансформації.
1931 рік - X. Даусон і Дж. Алловей підтвердили відкриття Ф. Гріффіта.
1934 рік - В. Стенлі, Ф. Боуден і Н. Пірі, досліджуючи рослинні віруси, дійшли висновку, що всі віруси містять нуклеїнову кислоту.
1934 рік - Т. Касперссон показав зв’язок молекул ДНК з хромосомами. 1934 рік - Хаммарстен і Т. Касперссон виявили, що молекули ДНК мають велику молекулярну масу (∞ 500000), яка переважає масу молекул білка. 1935 рік - у статті “Що таке ген?” М. Демерец передбачив, що спадковість пов’язана з молекулою ДНК.
1936 рік - А. М. Білозерський та І. І. Дубровська виділили ДНК у чистому вигляді із ядер клітин рослин.
1944 рік - О. Е. Айвері, К. Мак-Леод і М. Мак-Карті систематично вивчили роль різних речовин у клітині в явищах трансформації. Було доведено, що з явищем трансформації пов’язані молекули ДНК.
1948 рік - А. Буавен, Р. Вендрелі і К. Вендрелі показали зменшення кількості ДНК у статевих клітинах наполовину порівняно із звичайними клітинами.
1947-1950 роки - відкриття Е. Чаргаффом закономірностей нуклеотидного складу ДНК (правило Е. Чаргаффа).
1. Сума піримідинових нуклеотидів дорівнює сумі пуринових: пурини = піримідини.
2.Вміст тиміну дорівнює вмісту аденіну, а вміст гуаніну дорівнює вмісту цитозину: А = Т; Г = Ц.
3. Кількість 6-аміногруп дорівнює кількості 6-кетогруп: Г+Т = А+Ц.
4. Відношення А+Т= Г+Ц видоспецифічне. Для ДНК вищих рослин, тварин і деяких мікроорганізмів це відношення більше 1.
2 квітня 1953 року стаття Уотсона і Кріка під назвою “ Структура ДНК” була відіслана в редакцію журналу “ Нейчер ” - відкриття структури ДНК.
1959 рік - вперше одноланцюгову ДНК виявлено в найдрібніших фагів £ х 174, одноланцюгова ДНК має кільцеву форму. Дволанцюгова РНК вперше відкрита в реовірусів .
1961 рік - визначені тип і загальна природа генетичного коду (Ф. Крік, Л. Барнет, С. Бреннер, Р. Уотс-Тобін).
1961 рік - Мармур і Доті відкрили явище ренатурації ДНК, встановивши специфічність і точність реакцій гібридизації нуклеїнових кислот.
1966 рік - Ніренберг, Очоа і Корана розшифрували генетичний код.
1968 рік - X. Корана здійснив хімічний синтез гена.
1975-1977 роки - Сенгер і Баррел, а також Максам і Гілберт розробили методи швидкого визначення нуклеотидної послідовності ДНК.
Дезоксирибонуклеїнова кислота (ДНК)
1.У клітині знаходиться один вид ДНК.
2.Кількість ДНК протягом життя клітини стала.
3. ДНК знаходиться у клітині в ядрі, мітохондріях, пластидах зелених рослин. (Спостереження подвоєння центріолей привели деяких дослідників до уявлення, що центріолі так само, як мітохондрії і пластиди, належать до саморедуплікуючих компонентів цитоплазми, хоча прямих даних про наявність ДНК у складі центріолей ще немає).
4. “Будівельними блоками”, з яких побудована молекула ДНК, є нуклеотиди. Кожний нуклеотид складається з пуринової або піримідинової основи, з’єднаної з вуглеводом дезоксирибозою, яка з’єднана із залишком фосфорної кислоти. Пуринові основи - аденін і гуанін; піримідинові основи - цитозин і тимін. Кожна пуринова або піримідинова основа зв’язана своїм атомом 9-N або 1-N з атомом 1'-С дезоксирибози, а атом 5'-С дезоксирибози етерифікований фосфатом.
Гідроксильна група (ОН) в атомі 3'-С дезоксирибози одного нуклеотиду , може етерифікуватися фосфатом іншого нуклеотиду, і в результаті утворюється динуклеотид . Таким чином, приблизно від 70 до > 108 нуклеотидів ; зв’язуються в полінуклеотидний ланцюг. Осьовий скелет такої молекули складається із залишків фосфата і дезоксирибози, які чергуються, тоді : основи приєднані збоку. Всі пентозні дезоксирибози, залишки одного ланцюга, орієнтовані атомом 5'-С в одному напрямі, а атомом 3'-С - у протилежному. Таким чином, кожний ланцюг полярний: він має 5'-кінець (фосфатні кінець) і З'-кінець (гідроксильний кінець).
5. Молекули ДНК складаються приблизно з 2000 - 108 і більше мононуклеотидів . У кожній молекулі два полінуклеотидні ланцюги об’єднані один подвійний ланцюг. Основи розташовані парами одна проти іншої з’єднані водневими зв’язками. За стереохімічними причинами до утворені пар здатні тільки комплементарні (які підходять одне до іншого) основ аденін і тимін з’єднуються між собою двома, а гуанін і цитозин – трьома водневими зв’язками. Два ланцюги розташовуються антипаралельно: 5'- кінець одного ланцюга лежить навпроти 3'-кінця іншого ланцюга. Ланцюги комплементарні за основами: наприклад, послідовності 5'- А-ГЦ-Ц-Т-3' протистоїть у другому ланцюгу послідовність 3'-Т-Ц-Г-Г-А-5'. Внаслідок комплементарності основ у кожному подвійному ланцюгу кількість А дорівнює кількості Т, а кількість Г - кількості Ц. У цей самий час так зване
А+Т співвідношення ![]() видоспецифічне. Г+Ц
видоспецифічне. Г+Ц
У вірусів ланцюг ДНК теж подвійний, тільки в деяких фагів (вірус бактерій) є одноланцюгова ДНК.
6. Геометрія
подвійної спіралі така, що сусідні пари основ знаходяться одна від одної на
відстані 0,34 нм і повернуті на 36° навколо осі спіралі. На один виток спіралі
припадає , таким чином , 10 пар основ ![]() = 10 і крок спіралі дорівнює
3,4 нм. Діаметр подвійної спіралі ДНК дорівнює приблизно 20 нм. У подвійній
спіралі ДНК утворюються жолобки. Це пов’язане з тим що вуглеводнофосфатний
стовбур розташований далі від осі спіралі, ніж основи. В подвійній спіралі є
два жолобки - великий і малий.
= 10 і крок спіралі дорівнює
3,4 нм. Діаметр подвійної спіралі ДНК дорівнює приблизно 20 нм. У подвійній
спіралі ДНК утворюються жолобки. Це пов’язане з тим що вуглеводнофосфатний
стовбур розташований далі від осі спіралі, ніж основи. В подвійній спіралі є
два жолобки - великий і малий.
7. Рентгенограми волокон ДНК, одержані Франкліном і Уілкінсом між 1950 і 1953 роками, дали дуже важливу для побудови подвійної спіралі інформацію. Ідеалізована дифракційна картина має вигляд хреста із рефлексів (плям), який утворюється через регулярну структуру ДНК. Відстань між шаровими лініями відповідає періоду 3,4 нм , тобто кроку подвійної спіралі, сильний рефлекс (пляма) на 10-ій шаровій лінії - періоду 0,34 нм, тобто відстані між парами основ. Ці параметри відносяться до В-форми ДНК.
8. Стабільність подвійної спіралі зумовлена різними взаємодіями. Частково за неї відповідальні водневі зв’язки між основами. Але, напевно, більш важливу роль відіграє міжплощинна взаємодія - стекінг. При цьому забезпечуються не тільки вигідні вандерваальсові контакти між атомами, але і виникає додаткова стабілізація завдяки перекриванню П - орбіталей атомів контактуючих основ. Стабілізація здійснюється також за рахунок сприятливого гідрофобного ефекту, який виявляється в тому, що неполярні основи захищені від безпосереднього контакту з розчинником. Навпаки, вуглеводнофосфатний стовбур з його полярними групами і зарядженими атомами експонований, що також стабілізує структуру.
9. Поліморфізм ДНК - це здатність подвійної спіралі набирати різної конформації. Рентгеноструктурні дослідження кристалів полінуклеотидів (корот- сих полінуклеотидів, так званих олігонуклеотидів) виявили 3 основних типи структур:
А-, В-, Z - форми. В-ДНК - це стандартна уотсон - кріківська структура, в якій площини пар основ перпендикулярні до осі подвійної спіралі.
В А-ДНК площини пар основ повернуті приблизно на 20° від нормалі до осі правої спіралі. На виток спіралі тут припадає 11 пар основ. А-ДНК утворюється при висушуванні волокон В-ДНК. Z-ДНК - це ліва спіраль з 12 парами основ на виток. Буква Z вказує на зигзагоподібну форму вуглеводнофосфатного стовбура ДНК у цій формі. Площини основ приблизно перпендикулярні до осі спіралі. В клітині ДНК звичайно знаходиться в В-формі, але окремі її ділянки можуть знаходитися в А-,Z- або навіть в іншій конформації. (Утворення Z - форми, хрестоподібних інших структур стимулюється надспіралізацією ДНК).
10. Структура ДНК.
Молекула ДНК містить інформативні ділянки. В інформативних ділянках послідовність основ (первинна структура) являє собою матеріальний еквівалент генетичної інформації. Кожне повідомлення закодовано специфічною послідовністю із чотирьох знаків - А, Г, Ц, Т, подібно до того, як письмові повідомлення кодуються знаками (буквами) алфавіту або азбуки Морзе. Вторинна структура ДНК - це подвійна спіраль: два полінуклеотидних ланцюги закручені навколо загальної (спільної) уявної осі, й між ними утворюються два спіральні жолобки (борозенки) неоднакової глибини. Подвійна спіраль стабілізована водневими зв’язками і гідрофобними взаємодіями. В хромосомах подвійні спіралі утворюють тяжі з двома кінцями. Подвійні спіралі ДНК, які знаходяться в цитоплазматичних органелах (мітохондріях, пластидах) та в без’ядерних
(прокаріотичних) організмів, замкнуті в кільце, ЯКЕ може бути “зім’яте” в клубок.
У хромосомах подвійні спіралі з’єднані з білками (в основному іонними зв’язками) і утворюють разом з ними третинну структуру (суперспіраль у нуклеосомах, а також структури більш високого порядку).
11. Денатурація ДНК.
Структуру подвійної спіралі ДНК (або РНК), укріплену водневими зв’язками, можна зруйнувати нагріванням. Оскільки спарені ланцюги не зв’язані між собою ковалентно , після розриву всіх водневих зв’язків два полінуклеотидні ланцюги ДНК повністю розділяються.Процес розділення ланцюгів називають денатурацією або плавленням. Денатурація відбувається у вузькому інтервалі температур і відображається в кардинальній зміні багатьох фізичних властивостей ДНК. Особливо корисною виявилася зміна оптичної густини. Гетероциклічні кільця нуклеотидів поглинають світло в ультрафіолетовій : ділянці (з максимумом близько 260 мкм, характерним кожній основі). Але поглинання, характерне самій ДНК, майже на 40% менше, ніж поглинання суміші вільних нуклеотидів того самого складу. Це явище, яке називають гіпохромним ефектом , зумовлене взаємодією електронних систем основ у результаті їх стекінг-взаємодій при паралельному розташуванні в подвій: спіралі. Будь-яке відхилення від двоспірального стану відразу виявляється зміні величини цього ефекту. Іншими словами, відбувається зсув оптичної густини в бік значення, характерного для вільних основ.
Таким чином, за денатурацією ДНК можна спостерігати, досліджуючи її гіперхромність. Середню точку температурного діапазону, при якій прохододить розділення ланцюгів ДНК, називають точкою плавлення і позначають Тпл . Крива плавлення має завжди одну і ту саму форму, але її положення на температурній шкалі залежить від складу основ ДНК і умов, що використовуються для денатурації. Коли ДНК знаходиться в розчині приблизно в фізіологічних умовах, Тпл лежить у діапазоні 85-95 °С. Точне значення залежить від складу основ. Пари ГЦ, зв’язані трьома водневими зв’язками, мають велику вільну енергію утворення і виявляються більш тугоплавкими, ніж пари А-Т, що зв’язані двома водневими зв’язками. Залежність Тпл від складу основ лінійна. При кожному збільшенні вмісту Г-Ц- пар значення Тпл збільшується на 0,40 С .Так, ДНК, на 40% складена із Г-Ц (що типово, наприклад, для геному ссавців ) буде денатурувати при Тпл близько 87 °С у звичайних умовах, тоді як ДНК, містить 60% Г-Ц, за цих самих умов буде мати Тпл близько 95 °С. Вирішальний вплив на Тпл виявляє іонна сила розчину. Тпл зростає на 16,6 °С при кожному десятикратному збільшенні концентрації моновалентних катіонів. Частіше всього реакцію проводять у 0,12 М фосфатному буфері, що забезпечує концентрацію моновалентних іонів Nа +, рівну 0,18 М.
Значення Тпл сильно змінюється при добавленні в реакційну суміш таких речовин, як формамід , які дестабілізують водневі зв’язки. Їх присутність дозволяє знизити Тпл до 40 °С і цим самим запобігти додаткових пошкоджень ДНК (таких, як розрив ланцюгів), що викликаються підвищенням температури.
Надзвичайно корисна властивість денатурації ДНК - оборотність процесу. За певних умов два розділені комплементарні ланцюги можуть відновити подвійну спіраль. Це явище називають ренатурацією . Ренатурація залежить від специфічності спарювання основ між комплементарними ланцюгами, реакція відбувається у дві стадії. Спочатку короткі комплементарні послідовності двох ланцюгів випадково з’єднуються одна з одною і утворюють двоспіральну ділянку. Потім ділянка спарювання, подібно до замка блискавки, ширюється вздовж молекули і утворюється довга дволанцюгова структл Реконструкція подвійної спіралі завершується відновленням початкових в. тивостей, втрачених при денатурації ДНК. Швидкість ренатурації тим біль чим більша однорідність ДНК за нуклеотидним складом.
12. У 1950 році Е. Чаргафф і його співробітники, досліджуючи склад ДНК методом хроматографії, вперше встановили наступні основні закономірності кількісного складу азотистих основ у ДНК:
1. Сума пуринових основ (А+Г) дорівнює сумі піримідинових (Т+Ц).
2. Кількість основ з аміногрупами (А+Ц) дорівнює кількості основ з кетогрупами (Г+Т).
3. Молярні відношення пуринів до піримідинів близькі до 1 (А=Т і Г=Ц), тобто знаходяться в еквімолярних кількостях. Це положення має важливе значення при формуванні подвійної спіралі ДНК і відоме як правило еквівалентності.
Нуклеотидний склад ДНК у організмів
різних таксономічних груп специфічний і визначається співвідношенням ![]()
Це співвідношення називають коефіцієнтом
специфічності. За допомогою коефіцієнта специфічності був визначений
ступінь гетерогенності нуклеотидного складу ДНК у організмів різного
походження. Так, у вищих рослин і тварин відношення![]() коливається незначно і має
значення більше 1.
коливається незначно і має
значення більше 1.
Для мікроорганізмів коефіцієнт специфічності змінюється в широких межах - від 0,35 до 2,70. Разом з тим, соматичні клітини даного біологічного виду містять ДНК одного і того самого нуклеотидного складу, тобто можна сказати, що за вмістом ГЦ-пар основ ДНК одного виду ідентичні. Визначення гетерогенності нуклеотидного складу ДНК за коефіцієнтом специфічності ще не дає інформації про її біологічні властивості. Останнє зумовлено різною послідовністю окремих нуклеотидних ділянок у полінуклеотидному ланцюгу. Це значить, що генетична інформація у молекулах ДНК закодована в специфічній послідовності її мономерних одиниць. Таким чином, нуклеотидна послідовність має важливе значення для визначення тонкої структури геному. ДНК містить нуклеотидні послідовності, на значенні ініціації і термінації процесів синтезу ДНК (реплікація), синтезу РНК (транскрипція), синтезу білка (трансляція). Є нуклеотидні послідовності, які слугують для зв’язування специфічних активуючих і інгибуючих регуляторних молекул, а також нуклеотидні послідовності, що не несуть якої-небудь інформації. Існують такі модифіковані ділянки, які захищають молекулу від дії нуклеаз.
13. Передумовою для збереження наявної спадкової інформації в ряді послідовних поколінь клітин і організмів є ідентичне подвоєння, або реплікація генетичного матеріалу. Реплікація ДНК відбувається перед кожним нормальним діленням структур, що містять ДНК (ядер, пластид і мітохондрій) у еукаріот, перед кожним діленням бактеріальних клітин і при розмноженні ДНК- вірусів. У якості елементарних блоків для побудови нової ДНК у клітині синтезуються трифосфати чотирьох дезоксирибонуклеозидів - а Г, а Ц, а Т і а А.
Матрицею (шаблоном) для синтезу ДНК слугує наявна видо-специфічна дволанцюгова молекула ДНК. До обох ланцюгів прибудовуються комплементарні нуклеозидтрифосфати за принципом спарювання основ. За допомогою полімераз вони зв’язуються у новий нуклеотидний ланцюг, причому від кожного з них відщеплюється пірофосфат. Таким чином, на кожному ланцюгу старої молекули утворюється новий комплементарний ланцюг; реплікація відбувається, як кажуть, напівконсервативним способом. При редуплікації з одної молекули дволанцюгової ДНК утворюється дві молекули, ідентичні одна одній і вихідній молекулі. Це слугує передумовою збереження видоспецифічної спадкової інформації в ряді поколінь клітин і організмів, передумовою значної постійності видів.
Редуплікація бактеріальних геномів та інших кільцевих молекул ДНК починається у певній, генетично фіксованій “точці старту”. У хромосомах еукаріот є по декілька таких початкових точок. Редуплікація ДНК відбувається частинами. Репліковані часткові послідовності з 1000-2000 нуклеотидів називають фрагментом Оказакі . На обох старих ланцюгах новий поліпептидний ланцюг синтезується у напрямі 5' -> 3'. Починаючись точкою старту, реплікація здійснюється послідовно на окремих ділянках; при цьому в еукаріот і багатьох бактерій вона йде вздовж подвійної спіралі ДНК не тільки в одному напрямі, а і в протилежному. ДНКполімерази можуть приєднувати нуклеотиди не тільки вільного 3 '-ОН-кінця нуклеотиду, вже зв’язаного з старим ланцюгом ДНК. Тільки РНК-полімерази спроможні зв’язати перший нуклеотид з ДНК і почати новий полінуклеотидний ланцюг. Тому для того, щоб міг синтезуватися фрагмент Оказакі, спочатку повинен бути прибудований короткий відрізок РНК (як при транскрипції). Ця послідовність РНК із 1 - 10, рідше - близько 50 нуклеотидів, називається затравкою (або праймером) і синтезується РНК - полімеразою, або примазою. Від 3'-кінця затравки за допомогою ДНК - полімерази III починається синтез ДНК-фрагмента Оказакі, який продовжується до кінця даного фрагменту.
Наступний фрагмент Оказакі знову починається з РНК – затравки . ДНКполімераза І видаляє затравки, а пропуски заповнюються шляхом синтезу ланцюга ДНК, приєднаного до попереднього фрагмента Оказакі. Потім фермент лігаза зв’язує між собою синтезовані відрізки ДНК. Для реплікації подвійна спіраль ДНК розкручується ферментами. Реплікація на старому ланцюгу, що йде від точки старту в напрямі 3'—>5', може йти безперервно вздовж подвійної спіралі, яка розкривається подібно до застібки-блискавки. Реплікація на другому ланцюгу відбувається окремими фрагментами, оскільки на цьому ланцюгу новий ланцюг повинен синтезуватись у протилежному напрямі (відповідаючи також напрямку 3' > 5' старого ланцюга). Спарювання основ за участю ДНК- полімерази відбувається майже безпомилково. Перед зв’язуванням нового нуклеотида додатково перевіряється, чи правильним було попереднє спарювання. Якщо воно було помилковим, непотрібний нуклеотид видаляється завдяки 3'- 5' ендонуклеазній активності полімерази. При реплікації бактеріальних ДНК та інших кільцевих ДНКструктур у кінцевому підсумку одержуються дві ідентичні кільцеві молекули ДНК. В еукаріот різні реплікаційні ділянки хромосоми в кінці кінців об’єднуються, так що після завершення фази Б у кожній хромосомі знаходяться дві молекули дволанцюгової ДНК, які стають двома ідентичними хроматидами. Структура, здатна до реплікації (наприклад, хромосома, плазміда, вірусний геном), називається репліконом. Тільки структури з властивостями реплікона можуть зберігатися в ряді поколінь, тобто спадкуватися. ДНК здатна до реплікації завдяки наявності специфічної ділянки, яка може слугувати стартовим пунктом для реплікації. В еукаріот реплікація ДНК відбувається в інтерфазі. В бактерій реплікація ДНК починається в умовах, сприятливих для росту. Може відбутися декілька циклів реплікації один за одним. При цьому ті ділянки геному, з яких починається реплікація, можуть виявитися повтореними в клітині багаторазово. У фагів Т4 приблизно за 10 секунд при 14 °С зв’язується 1000 нуклеотидів.
14. Генетична інформація закодована в ДНК. Інформація, яка знаходиться в клітинному ядрі, являє собою генотип. ДНК, що міститься в одному наборі хромосом, називається геномом, а позаядерна ДНК (у мітохондріях, пластидах і основній речовині цитоплазми) - плазмоном. У бактерій ДНК у еквіваленті ядра являє собою геном, а позаядерна ДНК представлена у формі плазмід. Структури, які складаються із ДНК, що зустрічаються в основній речовині цитоплазми в еукаріот, так само як і в бактерій, називають плазмідами.
Генетична інформація в геномі бактерій і багатьох вірусів знаходиться в одномуєдиному безперервному полінуклеотидному ланцюгу. В еукаріот генетичний матеріал розподілений по хромосомах і в кожній хромосомі утворює один довгий нуклеотидний ланцюг. Полінуклеотидні нитки ДНК, які містяться в хромосомах еукаріот, в геномі бактерій і вірусів або плазмідах (у деяких вірусів - РНК), поділяються на функціональні відрізки, які називаються генами.
Розрізняють:
1.Структурні гени, в яких закодована інформація для синтезу ферментних і структурних білків.
2. Гени з інформацією для синтезу т-РНК (транспортні РНК).
3. Гени з інформацією для синтезу рибосомальної РНК (р-РНК).
4. Специфічні регуляторні ділянки, такі як промотори і оператори.
(Промотор - передуюча гену послідовність приблизно із 80 нуклеотидів, яку пізнає і з якою зв’язується фермент РНК-полімераза; оператор - це послідовність приблизно із 30 нуклеотидів, яку “впізнає” репресор. Ця послідовність має дзеркальну симетрію. Репресор являє собою алостеричний білок, який може приймати дві специфічні ділянки: одна для приєднання до оператора і одна для зв’язування ефектора. Приєднуючись до оператора, репресор блокує транскрипцію).
5. Розділяючі ділянки між генами (спейсери). (Спейсери - неінформативні відрізки ДНК різної довжини (інколи більше 20000 пар основ), які, напевно, мають значення для регулювання транскрипції сусіднього гена).
6. Ділянки з невідомою функцією (конститутивний гетерохроматин завжди гетерохроматичний. Він складається із послідовностей основ, які багаторазово повторюються, не інформативний (не містить генів) і тому завжди не активний стосовно транскрипції. Його можна бачити і під час ділення ядер. Найчастіше він зустрічається біля центромери, на кінцях хромосом (включаючи сателіти), а також поблизу організатора ядерця).
У 1977 році з’явилась можливість визначати послідовності нуклеотидів у ДНК. З тих пір визначені послідовності для деяких повних геномів і для багатьох частин геномів та зроблені висновки про їхні функції.
15. Під час трансляції нуклеотидна послідовність і-РНК переводиться в амінокислотну послідовність поліпептидного ланцюга. Генетичний код слугує ключем для переведення послідовності нуклеотидів у послідовність амінокислот. Як при перекладі з однієї мови на іншу необхідно знати слова, так і тут потрібно знати, як кожна амінокислота закодована в нуклеотидах. За період з 1961 по 1964 роки завдяки працям дослідницьких груп Ніренберг, і Очоа вдалось вияснити генетичний код. Результати цих праць виявились одним з найзначніших кроків у розумінні життєвих процесів. Ці результати можна коротко резюмувати наступним чином: - Генетичний код являє собою триплетний код. Триплет і-РНК одержав назву кодона.
-Генетичний код є виродженим кодом, тобто одній амінокислоті, як правило, відповідає більше ніж один кодон. У кодонах для одної амінокислоти перші два нуклеотиди найчастіше однакові, а третій - варіює.
- Нуклеотидна послідовність зраховується в одному напрямі підряд триплет за триплетом. Кодони не перекриваються.
- АУГ являє собою стартовий кодон.
- УАГ, УАА і УГА - кодони-термінатори.
- Генетично код універсальний, він єдиний для всіх організмів і вірусів
РНК (рибонуклеїнова кислота)
1. У клітині існує 3 основних типи РНК, які відрізняються локалізацією розмірами, нуклеотидним складом, структурою і функціональними властивостями: рибосомальна (р- РНК), транспортна (т -РНК) і матрична (м -РНК або інформаційна (і - РНК).
2. Кількість РНК протягом життя клітини змінюється.
3.РНК знаходиться в ядрі, цитоплазмі, рибосомах, мітохондріях, пластидах зелених рослин.
4.Будівельними блоками РНК є мононуклеотиди (або просто нуклеотиди). Кожний нуклеотид складається з одної пуринової чи піримідинової основи, з’єднаної з вуглеводом рибозою, яка з’єднана із залишком фосфорної кислоти. Азотистими основами РНК є: пуринові основи - аденін і гуанін: піримідинові основи - цитозин і урацил. Азотисті основи (тимін з ДНК урацил з РНК) відрізняються тільки метильною групою, яка присутня в тиміні . Таким чином, тимін слід було б назвати метилурацилом . Кожна пуринова основа (аденін, гуанін) і піримідинова основа (цитозин , урацил) зв’язана своїм атомом 9-N або 1-N з атомом 1 '-С-рибози. У свою чергу атом 5'-С-рибози етерифікований фосфатом. 5. Молекули РНК, як правило, одноланцюгові. В 1959 р. відкрито дволацюгову РНК у реовірусів . Гідроксильна група (ОН) атома 3'-С- рибози одного нуклеотиду може етерифікуватися фосфатом іншого нуклеотиду , і в результаті утворюється динуклеотид . Таким чином, приблизно від 70 до > 108 нуклеотидів зв’язуються в полінуклеотидний ланцюг. Осьовий скелет такої молекули складається із залишків фосфата і рибози, що чергуються, тоді як основи приєднані збоку.
Рибосомальна РНК (р-РНК).
Частка р-РНК складає до 90% усієї РНК клітини, вона характеризується метаболічною стабільністю. Це найбільші РНК, до складу їхніх молекул входить 3000-5000 нуклеотидів, відповідно їхня молекулярна маса становить 1000000-1500000. У прокаріот розрізняють 3 різних типи р -РНК, а в еукаріот розрізняють 4 різних типи р -РНК. РНК цього типу - локалізовані в рибосомах і беруть участь у специфічній взаємодії з рибосомними білками. Рибосомні РНК мають форму вторинної структури у вигляді коротких двоспіральних ділянок, з’єднаних зігнутим одиночним ланцюгом (Y або V- подібна форма). Білки рибосоми зв’язані переважно з однотяжевими ділянками молекули. Для р -РНК характерна наявність модифікованих основ, але в значно меншій кількості, ніж т -РНК.
Транспортна РНК (т-РНК).
Молекули т -РНК являють собою єдиний ланцюг, який складається з 70- 90 нуклеотидів, з молекулярною масою 23000-28000. У клітинній РНК транспортна РНК складає 10-20%. Молекули т -РНК володіють здатністю ковалентно зв’язуватись з певною амінокислотою і з’єднуватись через систему водневих зв’язків з одним з нуклеотидних триплетів молекули інформаційної РНК. Таким чином, т -РНК реалізує кодову відповідність між амінокислотою і відповідним її кодоном і- РНК. Для виконання адапторної функції т- РНК повинні мати цілком певну вторинну і третинну структури. Кожна молекула т- РНК володіє постійною вторинною структурою, має форму двомірного листка конюшини і складається зі спіральних ділянок, утворених нуклеотидами одного і того самого ланцюга, та розташованих між ними одноланцюгових петель. Кількість спіральних областей досягає половини молекули. Неспарені послідовності утворюють характерні структурні елементи (гілки), серед них можуть бути наступні типи гілок:
а) акцепторне стебло, на 3'-ОН кінці якого в більшості випадків розташований
триплет ЦУА он. До карбоксильної групи кінцевого аденозину за допомогою специфічного ферменту приєднується відповідна амінокислота;
б) псевдоуридилова петля, яка складається з 7 нуклеотидів ; вважають, що вона
використовується для зв’язування т- РНК з рибосомою;
в) додаткова петля - різна за розміром і складом у різних т- РНК;
г) антикодонова петля, яка складається з 7 нуклеотидів і містить групу з трьох
основ (антикодон); вона комплементарна триплету (кодону) у молекулі і- РНК;
д) дигідроуридилова петля, яка складається з 8-12 нуклеотидів і містить від
одного до чотирьох дигідроуридилових залишків; вважають, що вона використовується для зв’язування т- РНКзі специфічним активуючим ферментом.
Третинна укладка молекул т- РНК є досить компактною і має Г-подібну неправильну форму. У стабілізації третинної структури т- РНК беруть участь полівалентні катіони (Мg 2+, поліаміни ), а також водневі зв’язки між основами і фосфодіефірним стовбуром. Складна просторова укладка молекул т- РНК зумовлена множинними високоспецифічними взаємодіями як з білками, так і з іншими нуклеїновими кислотами (р- РНК). Транспортна РНК відрізняється від інших типів РНК високим вмістом модифікованих основ. Локалізація модифікованих нуклеотидів строго фіксована. Їх наявність у складі т- РНК зумовлює стійкість молекул до дії нуклеаз і, крім того, вони беруть участь у підтриманні певної структури, оскільки подібні основи не здатні до нормального спарювання і перешкоджають утворенню подвійної спіралі. Таким чином, наявність модифікованих основ у складі т- РНК зумовлює не тільки її структуру, а також і багато спеціальних функцій молекули т- РНК. У більшості клітин еукаріот міститься набір різних т- РНК (приблизно 60). Це означає, що для кожної амінокислоти є не менше, ніж по одній специфічній т- РНК. Виходячи з цього, т- РНК, що зв’язують одну і ту саму амінокислоту, називають ізоакцепторними. Кожний тип клітин у організмі відрізняється своїми співвідношеннями ізоакцепторних т- РНК.
Матрична (інформаційна) РНК (і-РНК, або м-РНК).
і- РНК містить генетичну інформацію про послідовність амінокислот для
основних ферментів та інших блоків, тобто слугує матрицею для біосинтезу поліпептидних ланцюгів. Частка і- РНК у клітині складає менше 5% загально: кількості РНК клітини. На відміну від р- РНК і т- РНК, і- РНК гетерогенна за розмірами, її молекулярна маса знаходиться в межах 25•103до 1•106; вміст нуклеотидів коливається від 300 до 3000. Наявність у клітині ланцюгів і- РНК змінної довжини відображає різноманітність молекулярних мас білків, синтез яких вони забезпечують. За своїм нуклеотидним складом і- РНК відповідає ДНК тієї самої клітини, тобто є комплементарною до одного з ланцюгів ДНК. Встановлено, що у послідовності нуклеотидів (первинна структура) і- РНК закладена інформація не тільки про структуру білка, але і про вторинну структуру самих молекул і- РНК . Вторинна структура і- РНК формується за рахунок взаємокомплементарних послідовностей, вміст яких у РНК різного походженню подібний і складає від 40 до 50%.
Третинна структура і- РНК формується головним чином за рахунок водневих зв’язків, гідрофобної взаємодії, геометричного і стеричного обмеження електричних сил. Інформаційна РНК являє собою метаболічно активну відносно нестабільну короткоживучу форму. Так, і- РНК мікроорганізмів характеризується швидким оновленням. Час її життя складає декілька хвилин. Разом з тим, для організмів, клітини яких містять істинні обмеження мембраною ядра, тривалість життя і- РНК може досягати багатьох годин навіть декількох діб. Стабільність і- РНК може визначатися різного роду модифікаціями її молекул.
Синтез білка. Транскрипція і трансляція. Генетичний код
ДНК бере участь у синтезі всіх білків, вона визначає їхню будову і функції. Але цілий ряд даних вказує на те, що сама ДНК безпосередньо не може бути матрицею в синтезі білків. У всіх клітинах, крім бактеріальних, майже всі ДНК знаходиться у хромосомах клітинного ядра, а в той самий час добре відомо, що синтез білка іде головним чином у цитоплазмі, де ДНК міститься у надзвичайно малих кількостях.
Значить, вже сам факт просторового розділення ДНК, яка знаходиться ядрі, і білків, синтезованих у цитоплазмі, вказує на існування якоїсь проміжної матриці, що переносить генетичну інформацію із ядра до цитоплазми - місця синтезу білків. У клітинах бактерій, які не мають ядра, ДНК і РНК просторово не розміщені, і наявність проміжної матриці для перенесення генетичної інформації на прикладі цих організмів так просто довести не можна. Але саме на бактеріях у експерименті Е. Волкіна та Л. Астрахана вперше було показано, що проміжною матрицею у процесі біосинтезу білка є РНК.
У цьому досліді ДНК фага дозволяли проникнути в клітини бактерій, а через деякий час, коли синтез клітинної РНК припинявся, в середовище вводили ортофосфорну кислоту, помічену радіоактивним фосфором 32Р. Виявилось, що РНК, яка утворюється у вихідній клітині, за складом основ була подібна з фаговою ДНК, на якій вона, очевидно, і синтезувалася. ДНК фага мала комплементарність основ: А=Т і Г=Ц, а РНК, знову утворена в клітинах бактерій, була подібна з нею за складом основ і виявила таку саму комплементарність: А=У і Г=Ц.
Прямі докази того, що безпосередньо на ДНК білок не синтезується і проміжною матрицею у цьому процесі, без сумніву, слугує РНК, були одержані у дослідах з використанням радіоавтографії. Точними дослідами встановлена пряма залежність між вмістом у клітинах РНК і кількістю синтезованого ними білка. Клітини, багаті РНК, синтезують більше білка, ніж клітини, які мають понижений її вміст.
Сукупність великого числа фактів і спостережень призвела до формулювання основного генетичного постулату матричної теорії спадковості, який схематично, зазвичай, виражається так:
ДНК (транскрипція) РНК (трансляція)
(реплікація) білок
Реплікація - процес самоподвоєння ДНК, у якому роль матриці відіграє сама молекула ДНК. Зігнута стрілка пояснює, що при реплікації молекули ДНК розмножуються шляхом самокопіювання. Транскрипція - перенесення
(переписування) інформації про нуклеотидну будову ДНК на РНК. Стрілка вказує, що РНК утворюється на ДНК- матрицях. Трансляція - процес, у якому матрицею для біосинтезу білка служить РНК. Вона визначає послідовність амінокислот у всіх білках. Стрілка вказує, що відбувається трансляція або переведення інформації про нуклеотидну будову РНК на амінокислотну будову білка. Значить, ДНК, входячи до складу ядра клітини, завдяки властивості самоподвоєння молекул зберігає свою кількісну постійність при поділі клітини, визначає структуру і регулює синтез утворених у клітині білків. Але молекули ДНК не є безпосередньо матрицями у самому процесі синтезу білка. Спочатку відбувається перенесення генетичної інформації про нуклеотидну будову ДНК на РНК. Потім остання сама стає матрицею і у відповідності з інформацією, одержаною від ДНК, визначає послідовність з’єднання амінокислот у білковій молекулі.
Тепер, коли нам у загальних рисах відома пануюча у молекулярній генетиці теорія спадковості, розглянемо, як відбувається транскрипція генетичної інформації. Вона здійснюється шляхом синтезу і- РНК на ДНК- матриці. Назву інформаційної ця РНК одержала тому, що вона, проникаючи через пори ядерної оболонки, несе до цитоплазми (місця синтезу білка) інформацію про порядок чергування нуклеотидів у молекулі ДНК. Будується молекула і- РНК на одному з ланцюжків молекули ДНК- матриці під час її роздвоєння за участю спеціального ферменту – РНК – полімерази . Спарювання нуклеотидів іде за принципом компліментарності : послідовність нуклеотидів у молекулі і- РНК визначається їхньою послідовністю у ланцюжку ДНК, при цьому гуанілова кислота з’єднується з цитидиловою, тимідилова - з аденіловою , а аденілова кислота ДНК не з тимідиловою, як це буває при реплікації ДНК, а з уридиловою кислотою. Одна молекула і -РНК, як правило, несе інформацію про будову одного поліпептидного ланцюга. Тільки-но закінчується побудова на ДНК- матриці ланцюга і- РНК, вона відразу ж переходить у цитоплазму і прикріплюється там до однієї з рибосом.
Слідом за цим починається синтез білка.
Але насамперед необхідно ознайомитися з генетичним кодом. Будь-які відмінності у структурному і кількісному складах білків змінюють їхні властивості, тому одне з головних питань спадковості полягає у виясненні того, як генетична інформація, записана у хімічній структурі молекул ДНК, передається в процесі біосинтезу специфічних білків, яким чином вона перетворюється у всі речові і функціональні ознаки і властивості, якими визначаються особливості наступних поколінь клітин і організмів. Проблема взаємовідношення білків і нуклеїнових кислот - центральна у вченні про спадковість. Білки - біологічні полімери їхні макромолекули складаються всього з 20 мономерів - амінокислот. Амінокислоти можуть входити у молекули білків у неоднакових кількостях, по-різному в них з’єднуватися і чергуватися між собою і по-різному розташовуватись у просторі. Таким чином, не дивлячись на велику різноманітність білків, вони відрізняються один від одного у своїй первинній структурі тільки порядком розташування амінокислот. 20 амінокислот можуть утворити 1024комбінацій. Оскільки будь-які відмінності в ознаках зводяться до відмінностей у білках, то зрозуміло, ще така кількість білків створює практично нескінченну різноманітність ознак і властивостей організмів. Вона може забезпечити продовження еволюції життя на Землі не менше ніж до 10 млрд. років. Відмінності хоча б в одній амінокислоті достатньо, щоб змінити властивість білка, а отже, і ознаку організму. Наприклад, заміна у білковій молекулі гемоглобіну, яка складається приблизно із 600 амінокислот, одної електрично зарядженої глутамінової кислоти на електрично нейтральну амінокислоту валін веде до важкого малокрів’я - хвороби, що дістала назву серповидноклітинної анемії. Червоні кров’яні тільця (еритроцити) у таких хворих набувають форми півмісяця втрачають електричний заряд і не здатні тому зв’язувати молекули кисню. Діти, хворі на серповидноклітинну анемію, вмирають у ранньому віці. Подібні хвороби, що виникають у результаті мутацій, які зачіпають молекулярна будову білків, називаються молекулярними хворобами.
Яким чином молекулярна будова ДНК визначає біосинтез різних білків ДНК такий самий полімер, як і білок. Ланцюг ДНК також побудований із мономерних ланок, що чергуються. Але якщо у білків їх - 20, то у ДНК - всього 4. Якщо відомі нам нуклеотиди А, Г, Ц, Т, оскільки вуглевод і фосфор на кислота в усіх нуклеотидах однакові, різняться між собою тільки азотистими основами, то відмінності між ДНК зводяться лише до порядку розміщення азотистих основ. їхня послідовність у молекулі ДНК визначає послідовність амінокислот у молекулі білка. Отже, форми і функції усіх організмів, їхні індивідуальні і видові відмінності визначаються комбінацією чотирьох азотистих основ молекули ДНК. Послідовність розташування азотистих основ у ДНК, що визначає розміщення амінокислот у синтезованому білку, називається генетичним кодом, або кодом спадковості.
Ідея генетичного коду допускає порівняння азотистих основ, що входять у ДНК, з буквами алфавіту (за аналогією між записом і передачею будь-яких слів та наявних у них відомостей (інформації) через різне поєднання букв) і визначенням, “ записом ” структури і функцій білкових молекул шляхом різного поєднання чотирьох азотистих основ у молекулі ДНК. Саме у цьому розумінні говорять, що спадкова інформація “ записана ” у молекулах ДНК.
Оскільки одна і та сама спадкова інформація “ записана ” в нуклеїнових кислотах чотирма знаками (азотистими основами), а в білках - двадцятьма знаками (амінокислотами), проблема генетичного коду зводиться до встановлення відповідності між ними. Робота по розшифруванню генетичного коду вимагала прикладення зусиль вчених різних спеціальностей: генетиків, фізиків, хіміків, математиків. Особливо значну роль у вирішенні цієї проблеми зіграли дослідження фізика Г. Гамова і генетика Ф. Кріка — одного з авторів моделі будови молекули ДНК.
Для розшифрування генетичного коду насамперед необхідно було вияснити, яке мінімальне число нуклеотидів може визначати (кодувати) утворення однієї амінокислоти. Якщо б кожна із 20 амінокислот кодувалась одною основою, то ДНК повинна була б мати 20 різних основ, фактично ж їх тільки 4. Очевидно, поєднання двох нуклеотидів також недостатньо для кодування 20 амінокислот. Воно може кодувати лише 16 амінокислот (42=16 поєднань). Поєднання ж з 3 нуклеотидів дає 64 комбінації (43=64 поєднання) і, отже, здатне кодувати більше ніж достатнє число амінокислот для утворення будь-яких білків. Таке поєднання трьох нуклеотидів називається триплетним кодом. У триплетному коді амінокислоти кодуються трійками основ (наприклад, УУУ, ЦГЦ, АЦА і т. д.). Ділянка ланцюга ДНК із трьох нуклеотидів, який визначає включення у білкову молекулу строго певної амінокислоти, називається кодоном.
Після ґрунтування принципу генетичного коду необхідно було експериментальним шляхом встановити, які конкретні триплети кодують кожну із 20 амінокислот. Початок вирішення цього складного завдання було покладено у дослідах американських біохіміків М. Ніренберга та Дж. Маттеї. У 1961 році на V Міжнародному біохімічному конгресі у Москві М. Ніренберг доповів про відкриття триплету, який кодує синтез амінокислоти фенілаланіну. Ніренберг і Маттеї використали у своїх дослідах найпростішу РНК - поліуридилову кислоту. Ця синтетична РНК побудована тільки з уридилових нуклеотидів:
ФР - ФР - ФР - ФР
І І І І
У У У У
Поліуридилову кислоту добавляли в якості матриці у виділений із клітини розчин, що містив рибосоми. Осад, що при цьому випав, виявився білком. Але це був незвичайний білок - поліфенілаланін. Хімічний аналіз показав, що в білковий поліпептидний ланцюг поліфенілаланіну зв’язувались молекули тільки однієї єдиної амінокислоти - фенілаланіну. В розчині були всі 20 амінокислот, але поліуридилова РНК вибрала тільки фенілаланін. Якщо виходити з цього, то амінокислотний код - триплетний, таким кодом для фенілаланіну, очевидно, є триплет, до складу якого входять 3 уридилові кислоти. Це і було експериментально доведено: дійсно, молекула РНК, яка складалася з триплетів урацилу (УУУ), кодує амінокислоту фенілаланін. Триплет УУУ був першим знаком генетичного коду, який вдалося розшифрувати. Потім було проведено багато дослідів, коли у безклітинну систему добавлялись інші синтетичні РНК і робився аналіз одержаних результатів. При добавлянні поліцитидилової РНК синтезувалась амінокислота пролін. Її кодоном виявився триплет ЦЦЦ. Якщо брали поліуридилову кислоту і добавляли поліаденіл (полі-А), то отримували білок, який складався головним чином із фенілаланіну, та в нього входив ще й ізолейцин. За співвідношенням полі-У та полі-А був встановлений код для ізолейцину - УУА. Так, послідовно комбінуючи в синтетичних РНК по три основи з чотирьох, у лабораторіях М. Ніренберга та С. Очоа в
1962 році був розшифрований склад нуклеотидних триплетів для всіх 20 амінокислот, що входять до складу білкових молекул.
Уявлення про триплетний код було підтверджено багато чисельними біохімічними і прямими генетичними експериментами, обґрунтованими на порівняльному аналізі мутацій. Отже, код триплетний, тобто визначення послідовності амінокислот у молекулі білка визначається шляхом специфічного поєднання трійок нуклеотидів у ланцюгу ДНК. Коли були розшифровані триплети для всіх амінокислот, що входять у молекули білка, виявилося, що більшість з них кодується не одним, а двома, трьома, навіть чотирма різними триплетами. Наприклад, метіонін кодується одним триплетом (АУГ), лізин — двома (ААА і
ААГ), ізолейцин - трьома (АУУ, АУЦ, АУА), серин - чотирма триплетами (УЦУ, УЦЦ, УЦА, УЦГ). У зв’язку з цим говорять про виродженість коду і код називають виродженим. Під виродженістю коду розуміють можливість включення в білкову молекулу одної амінокислоти декількома триплетами. Триплети не перекривають один одного, один триплет не може входити до складу інших, кожний з них самостійно кодує свою амінокислоту. Тому в поліпептидному ланцюгу поряд можуть знаходитися будь - які дві амінокислоти і можливі які завгодно їх поєднання. Код не має “ розділових знаків ”. На відміну від писемного коду, це, як кажуть, код без коми. Триплети нічим не розділені між собою. Списування коду відбувається лінійно, починаючи з якоїсь певно фіксованої в ланцюгу ДНК точки (початку гена), і йде тільки в одному напрямі. Стартовим сигналом до початку синтезу білка слугує розташований на і -РНК кодон АУГ, який кодує метіонін (мет) (іноді цей кодон для валіну – ГУЦ ). У поліпептидному ланцюгу, що росте, першим амінокислотним залишком завжди буде або метіонін, або валін. Тоді виникає закономірне питання: яким чином клітина відрізняє стартовий сигнал від кодонів АУГ або ГУЦ , розташованих у середині молекули і- РНК? Ця проблема вирішується за допомогою модифікованої форми метіоніну або валіну і спеціальної ініціюючої т -РНК. Формілметіонін (fмет) і є тою модифікованою формою метіоніну, з якої починається синтез білка. Він приєднується до молекул т -РНК певного типу (т -РНК,), ЯКІ відрізняються від T –PHK f мет , за допомогою яких метіонін включається в серединну частину поліпептидного ланцюга. І т -РНК, і т – РНКfмет пізнають кодон АУГ, але лише т -РНКг здатне приєднатися до стартового кодону АУГ. Випадання або вставка якого-небудь нуклеотиду в молекулі ДНК змінює склад і послідовність чергування триплетів у всьому їхньому ланцюгу. Ми знаємо, що кожна і -РНК синтезує один-єдиний білок. Але щоб це було можливим, повинно бути “ заборонено ” списування зі зсувом на одну або дві основи. В іншому випадку списування встановленого запису порушується. Неважко побачити, що випадання або вставка в триплеті будь-якого нуклеотиду викликає помилки у кодуванні, виникають “ безглузді ” триплети, склад їх змінюється, так що утворення запрограмованого білка стає неможливим.
На основі робіт багатьох вчених у середині 50-х років була висунута матрична теорія синтезу білка. Згідно цієї теорії, синтез білка - дуже складний багатоступінчастий процес. У ньому беруть участь ДНК, різні види РНК і різноманітні ферменти. Кожний білок синтезується на своїй особливій матриці і для цього потрібна своя особлива і -РНК. Одна молекула і -РНК транскрибує послідовність нуклеотидів з відрізка ДНК, рівного одному гену, і переносить цю інформацію на послідовність розташування амінокислот у поліпептидному ланцюгу одного білка. Інформаційна РНК, проникаючи з ядра у цитоплазму і прикріплюючись до рибосом, починає діяти стосовно білків як матриця.
Процес синтезу білка складається з 4 послідовних стадій, або етапів.
І етап - активування амінокислот, у результаті чого підвищується їхня активність і вони легше взаємодіють одна з одною при з’єднанні в поліпептидний ланцюг. У цитоплазмі завжди є набір амінокислот, необхідних для синтезу потрібних клітині білків, але перш ніж взяти участь у цьому процесі вони повинні бути активовані. Активування амінокислоти відбувається при її взаємодії з АТФ. Виникає сполука, у якій весь запас енергії АТФ переходить на амінокислоту, яка стає при цьому більш активною. Зв’язування АТФ з амінокислотою каталізує особливий фермент, який називається амінацил – РНК -синтетазою.
ІІ етап пов’язаний з перенесенням або транспортуванням активованих амінокислот до рибосом. Цю функцію виконує транспортна РНК (т -РНК). Молекула т -РНК порівняно з молекулою і -РНК невелика, вона містить усього 70-80 нуклеотидів. її полінуклеотидний ланцюг приблизно посередині перегинається, і дві половини спірально закручуються між собою. На одному кінці молекули т -РНК повинні бути основи, комплементарні відповідній ділянці (кодону) в ланцюзі і- РНК, а на другому кінці - здатні “ пізнавати ” певну амінокислоту.
Кінець, до якого приєднується амінокислота, у всіх т -РНК має однакові нуклеотиди - ЦЦА. Для кожної амінокислоти існує своя особлива т -РНК. Значить, повинно існувати не менше 20 різновидностей т -РНК, по одній для кожної із 20 амінокислот, з яких будуються різні білки. Кожна з молекул т -РНК приєднується до “ своєї ” активованої амінокислоти. Значить, т -РНК виконує у процесі синтезу білка роль адаптера, тобто речовини, яка доставляє і прикладає активовані амінокислоти до відповідних триплетів і –РНК -матриці.
ІІІ етап починається шикуванням амінокислот у порядку, визначеному чергуванням нуклеотидів ДНК на і –РНК -матриці і закінчується замиканням пептидних зв’язків у молекулі білка. Відбувається цей процес у рибосомах за участю фермента пептидполімерази. Рибосоми побудовані з білка і РНК. Ця РНК називається рибосомною (р -РНК). Функції р -РНК поки що точно не вияснені. Вважають, що неспарені основи р -РНК беруть участь у зв’язуванні т -РНК та і -РНК.
Рибосоми, зв’язані між собою в групи або агрегати, названі полісомами.
Електронно-мікроскопними дослідженнями встановлено, що у полісомах рибосоми з’єднані між собою ниткою рибонуклеїнової кислоти діаметром 0,001-0,0015 мкм. Число рибосом, що входять до полісоми, визначається довжиною молекулярного ланцюга і -РНК. Прикріпившись на кінці нитки і -РНК, рибосома, до якої т -РНК увесь час доставляють активовані амінокислоти, починає синтез поліпептидного ланцюга. Пересуваючись у одному певному напрямі, вона злічує по 3 нуклеотиди і добавляє до поліпептидного ланцюга, що росте, по одній амінокислоті. Досягнувши іншого кінця ланцюжка і -РНК, рибосома відділяється, і в розчин виходить нова синтезована молекула білка.
На полісомі одночасно синтезуються декілька поліпептидних ланцюгів одного і того самого білка. Різниця між ними лише у кількості зібраних амінокислот. Збірка білкових молекул на полісомі нагадує роботу стрічки конвеєра. Молекулярна швидкість трансляції і транскрипції велика - близько 1000 триплетів і -РНК за хвилину на одну рибосому, а всього за хвилину, наприклад, клітина Eschtrichia coli включає у білки близько 15 х 106 амінокислот. Деякі локуси транскрибуються протягом клітинного циклу більше 1000 разів.
ІVетап. У цей час лінійна молекула поліпептидного ланцюга набуває об’ємної структури. Під впливом водневих зв’язків, що виникають, поліпеп - тидний ланцюг скручується у спіраль, і білкова молекула набуває біологічно активної конфігурації.
Висновок. Провідна роль у синтезі білка належить ДНК. Залежно від розташування кодуючих триплетів вздовж її ланцюга на ній синтезується молекула і
-РНК, яка реалізує цю інформацію, розташовуючи, відповідно до будови, амінокислоти в синтезованій молекулі білка. Таким чином, спадкова інформація, всі ознаки і властивості організму зберігаються в молекулярній структурі ДНК, а реалізується спадковість у процесі біосинтезу білка.
Регуляція біосинтезу білка
Відповідно до інформації, закладеної в структурі ДНК, організм у процесі росту і розвитку синтезує всі необхідні йому білки. Всі соматичні клітини будь-якого вищого організму, як би вони не були диференційовані, містять однакову кількість ДНК одного і того самого типу і, значить, несуть одну і ту саму програму біосинтезу білків. Але різні види клітин відрізняються одна від одної за кількістю і типами білків, так само як і в одній і тій самій клітині у різний час синтез білків іде з неоднаковою швидкістю. Значить, у клітині повинен існувати якийсь дуже досконалий механізм, що забезпечує вибірковий синтез необхідних їй білків і потрібну в цей чи інший момент їх кількість. Спочатку у мікроорганізмів, а потім і в клітинах вищих рослин було виявлено
пристосоване регулювання синтезу ферментів. Виявилося, що клітини синтезують не всі білки, які вони потенційно здатні утворювати. Щоб у клітині почався синтез певного фермента, в неї з оточуючого середовища повинна проникнути якась речовина, здатна індукувати цей процес. Така речовина називаєся індуктором. Дуже часто ним буває природний субстрат ферменту . Наприклад, якщо в середовище з мікробними клітинами, які ростуть на глюкозі добавити інший вид цукру - лактозу, то вона ними буде зброджуватися так само, як і цукор. У цьому випадку лактоза виявилася тим самим специфічним субстратом-індуктором, який індукував утворення в мікробних клітинах новового фермента. Найцікавішим у цьому досліді є те, що новий фермент утворився в клітині тільки тоді, коли він їй необхідний. Але синтез фермента може не тільки пристосовано індукуватися, але й подавлятися, репресуватися. Подавлення синтезу ферменту відбувається тоді, концентрація якої-небудь речовини, яка виробляється клітиною, перевищує певний рівень. Часто таким репресором слугує яка-небудь амінокислота високої концентрації, токсична для клітини. Така амінокислота - репресор виключить синтез саме тих ферментів, які її синтезують. Наприклад, бактерії Eschtrichia coli при надлишку в середовищі триптофану перестають утворювати триптофан-синтетазу - фермент, який синтезує цю амінокислоту. Значить, з ферментів у клітині регулюється механізмами індукції та репресії.
Логічно було б передбачити, що індукція і репресія синтезу білків, як і які інші процеси клітинного метаболізму, знаходяться під контролем Це підтверджувалося даними вивчення у бактерій деяких біохімічних мутацій. Були виявлені мутації, що порушують механізм індукції або репресії. Наприклад, може відбутися мутація, внаслідок якої клітина починає безперервно синтезувати фермент незалежно від присутності або відсутності індуктора. Були виявлені мутантні штами кишкової палички, які синтезуваіли галактозидазу як у присутності, так і за відсутності індуктора — молочного цукру лактози.
Аналогічно мутація може вивести синтез ферменту з-під контролю репресора. При цьому клітина буде продовжувати утворювати фермент і тоді, в ньому немає потреби і продукт діяльності цього фермента є у надлишку. Особливість названих мутацій полягає у тому, що вони зачіпають не сам процес синтезу ферментів, а лише його регуляцію. Виходячи з цих фактів, внаслідок точних генетичних і біохімічних експериментів, французькі мікробіологи і генетики Ф. Жакоб і Ж.
Моно створили в 1961 році цілісну загальну теорію регуляції білкового синтезу. Розберемо основні засади цієї теорії й обґрунтовану на ній принципову механізму генетичного контролю синтезу ферментів у бактерій. Всі гени знаходяться у великій молекулі ДНК, яка самовідтворюється. Кожний з них собою невелику ділянку такої молекули. Але за своїми функціями гени неоднакові. Одні з них несуть інформацію про послідовність амінокислот у білковій молекулі, тобто визначають її структуру, інші регулюють активність їх і цим контролюють процес поступання інформації від ДНК до і -РНК. а група генів одержала назву структурних, друга - регуляторних. Структурні гени, які контролюють синтез ферментів, у якому-небудь одному ланцюгу реакцій, розташовані, зазвичай, поряд один з одним. Вони складають єдиний блок, названий опероном, і здійснюють послідовні етапи синтезу одного ферменту, працюючи погоджено, як один елемент. Згідно моделі будови хромосоми, запропонованої Ф. Кріком, структурна (інформативна) зона оперону, яка несе інформацію для синтезу білків, розташована в міждисковій частині хромосоми, регуляторна (акцепторна) її частина входить до складу дисків. Гени в опероні або всі активні, або всі пасивні. Гени одного оперона здійснюють всі поступальні (одна за одною) реакції синтезу кінцевого продукту. Тому синтезуються або всі ферменти у ланцюзі реакції, або не синтезується ні один з них. Вся група генів одного оперона включається в процес синтезу і виключається з нього одночасно. Включення і виключення структурних генів складає сутність всього процесу регуляції. Функції виключення і включення виконує особлива ділянка молекули ДНК - ген-оператор, розташований на самому початку оперона . Ген - оператор до тих пір, поки, до нього не приєднається молекула репресора, знаходиться у виключеному стані. Тільки-но репресор зв’язується з геном - опероном весь оперон виключається і його гени стають неактивними. Якщо репресор немає, структурні гени включаються і йде синтез молекул РНК, які несуть j цитоплазму інформацію для синтезу всього набору ферментів, що виробляють ся цим опероном.
Репресор - це речовина білкової природи. Він синтезується геном, розташованим на якійсь відстані від оперона. Цей ген називається геномрегулятором. Ген-регулятор безперервно посилає до цитоплазми і -РНК, яка містить інформацію для синтезу білків -репресорів. Таким чином, функція гена-регулятора полягає у керуванні синтезом молекул репресора, які потім з’єднуються з оператором і впливають на механізм включення структурних генів oneрона. Робота гена-регулятора, що виробляє молекули репресора, направляється і контролюється цитоплазмою клітини і залежить від зовнішніх умов.
Розглянемо, як здійснюється механізм регуляції. Поки кінцевий продукт утворюється у потрібній клітині кількості, репресор знаходиться в не активному стані, ген-оператор - “ вимкнений ” і структурні гени працюють Тільки-но кінцевий продукт починає вироблятися в кількості, більшій, ніж це потрібно клітині у даний момент, він вступає в реакцію з репресором, який активується і, зв’язуючись з геном-оператором, вимикає роботу всієї системи. Але коли у клітині знову виникає необхідність у біохімічній реакції внаслідок якої виробляється кінцевий продукт, дія репресора знімається . Відбувається це через індукцію. Індуктором зазвичай слугує та речовина яка переробляється за участю даного фермента, тобто є його субстратом. Молекули цієї речовини, з’єднуючись з репресором, цим самим одночасно звільняють ген-оператор, який вмикає роботу структурних генів, і синтез не обхідного продукту продовжується. Сигнал на увімкнення в роботу оперон дає речовина, вихідна в біологічній реакції, яка йде за участю синтезованого фермента , а сигнал на його вимикання поступає від речовини, яка утворюється в результаті цієї самої реакції. Дія такої двосторонньої системи сигналізації ґрунтується на тому, що молекули репресора володіють властивістю з’єднуватись і з геном-оператором, і з молекулами індуктора. Механізм геї ної регуляції синтезу ферментів добре вивчений у бактерій, що розщеплюють молочний цукор (лактозу) за допомогою фермента галактозидази, який вони виробляють. Мікробна клітина поглинає з оточуючого середовища лактозу і розщеплює її за допомогою фермента галактозидази. Якщо у розчині лактози немає, фермент для її розщеплення не потрібний, репресор, що виробляється геном-регулятором, вступає у взаємодію з геном-оператором і “ запирає ” його. Цим самим припиняється робота структурних генів, на яких синтезувалась і -РНК, яка слугує матрицею для вироблення фермента галактози. Тільки-но в оточуючому клітину середовищі з’являється лактоза, вона, діючи як індуктор, вступає у взаємодію з репресором і зв’язує його. Одночасно з цим ген-оператор звільняється від репресора і включає в роботу структурні гени: починається синтез і -РНК і утворення фермента галактозидази. Нові порції репресора, які виробляються геном-регулятором, продовжують зв’язуватися лактозою. Поки в оточуючому клітину середовищі є лактоза, весь час утворюється фермент, що її розщеплює. При втраті всіх запасів лактози репресор звільняється і вимикає роботу структурних генів. Утворення непотрібного більше клітині ферменту галактозидази припиняється. Як бачимо, генетична система клітини, використовуючи механізми індукції та репресії, може приймати сигнали про необхідність початку і закінчення синтезу того чи іншого фермента і здійснювати цей процес із заданою швидкістю. Таким чином, механізм регуляції білкового синтезу - це досконала здатна до самонастроювання та саморегуляції біокібернетична система, що ґрунтується на принципі дії зворотного зв’язку. Поступання керівної інформації від ДНК на синтез певного фермента регулюється потоком зворотної інформації про вироблену кількість цього ферменту і потреби у ньому клітини в кожний даний період.
Приклади розв’язання задач з молекулярної біології .
Задача 1. Дано ряд нуклеотидів: А (аденін), Т (тимін), У (урацил), Ц (цитозин), Г (гуанін). Визначте, які входять до складу ДНК , які до складу РНК.
Розв'язок.
У нуклеїнових кислотах тільки 4 азотистих основи. До складу ДНК і РНК входять наступні азотисті основи :
ДНК - А (аденін) , Т (тимін) , Г (гуанін) , Ц (цитозин) .
РНК - А (аденін) , У (урацил) , Г (гуанін) , Ц (цитозин) .
Видатний учений Е. Чаргафф визначив, що ці нуклеотиди в нуклеїнових кислотах розташовуються комплементарно, причому загальна кількість пуринів дорівнює загальній кількості піримідинів: (А + Т) + (Г + Ц) = 100% в ДНК ; (А + У) + (Г + Ц) = 100 % в РНК .
Задача 2. На фрагменті одного ланцюга ДНК нуклеотиди розташовані в послідовності:
А - А - Г - Т - Ц - Т - А - Ц - Г - А - Т - Г. Запишіть схему структури дволанцюгової молекули ДНК; поясніть, якою властивістю ДНК при цьому ви керувалися; яка довжина даного фрагмента ДНК. Примітка. Кожен нуклеотид займає 0,34 нм за довжиною ланцюга ДНК.
Розв'язок.
Ділянка одного фрагмента дволанцюгової молекули ДНК виглядає так:
А - А - Г - Т - Ц - Т - А - Ц - Г - А - Т - Г
Т - Т - Ц - А - Г - А - Т - Г - Ц - Т - А - Ц
Дволанцюгова молекула ДНК будується за принципом комплементарності (доповнення). Визначаємо довжину даного фрагмента ДНК.
До фрагменту входить 12 нуклеотидів, довжина кожного - 0,34 нм: 0,34 · 12 = 4,08 нм.
Задача 3. На фрагменті одного ланцюга ДНК нуклеотиди розташовані в такій послідовності: А - А - Г - Т - Ц - Т - А - Ц - Г - Т - А - Г.
Визначте схему структури дволанцюгової молекули ДНК, підрахуйте відсотковий склад нуклеотидів в цьому фрагменті.
Розв'язок.
Структуру дволанцюгової молекули ДНК визначаємо за принципом комплементарності :
А - А - Г - Т - Ц - Т - А - Ц - Г - Т - А - Г
Т - Т - Ц - А - Г - А - Т - Г - Ц - А - Т - Ц
Для визначення відсоткового складу нуклеотидів треба використовувати співвідношення:
(А + Т ) + ( Г + Ц) = 100%
Підрахуємо загальну кількість нуклеотидів:
А = 7 ; Т = 7 ; Г = 5 ; Ц = 5. Всього 24 . Визначимо, скільки відсотків припадає на один нуклеотид : 24/ 100 = 1 / х х = 100 · 1: 24 = 4, 17
Тепер можна розрахувати і кількість нуклеотидів:
24 / 100 = 7 / х ;
х = 100 · 7 : 24 = 29,17 (А і Т);
24 / 100 = 5 / х ; х = 100 · 5 : 24 = 20,83 (Г і Ц ) .
Можна визначити й іншим способом:
7 · 4,17 = 29,17 (А і Т); 5 · 4,17 = 20,83 (Г і Ц ).
Задача 4. Довжина фрагмента молекули ДНК дорівнює 20,4 нм. Скільки нуклеотидів в цьому фрагменті ?
Розв'язок.
У всіх організмів, крім деяких вірусів, молекула ДНК дволанцюгова, а розмір одного нуклеотиду становить 0,34 нм.
Визначаємо кількість нуклеотидів в цьому фрагменті: 20,4 : 0,34 = 60 (нуклеотидів).
Задача 5. Частковий і -РНК гена інсуліну має наступний склад: УУУ - ГУУ - ГАУ - ЦАА - ЦАЦ - УУА - УГУ - ГГГ - УЦА - ЦАЦ. Визначте співвідношення (А + Т) :
(Г + Ц) у фрагменті названого гена.
Розв'язок.
і -РНК - УУУ - ГУУ - ГАУ - ЦАА - ЦАЦ - УУА - УГУ - ГГГ - УЦА - ЦАЦ ДНКл.ц. - ААА - ЦАА - ЦТА - ГТТ - ГТГ - ААТ - АЦА - ЦЦЦ - АГТ - ГТГ ДНКп.ц. - ТТТ - ГТТ - ГАТ - ЦАА - ЦАЦ - ТТА - ТГТ - ГГГ - ТЦА - ЦАЦ Підрахуємо кількість нуклеотидів в ДНК: А = 18 ; Т = 18 ; Г = 12 ; Ц = 12 .
(А + Т ) : ( Г + Ц) = (18 + 18 ) : ( 12 + 12) = 36:24.
Задача 6. У молекулі ДНК виявлено 880 гуанілова нуклеотидів, які складають 22 % від загального числа нуклеотидів в цій ДНК.
Визначте: а ) скільки інших нуклеотидів в цій ДНК? б) яка довжина цього фрагмента?
Розв'язок.
1) І варіант розв'язку - Σ (Г) = Σ (Ц) = 880 (це 22%);
На частку інших нуклеотидів припадає: 100 % - (22% + 22%) = 56 % , тобто по 28 %; Для обчислення кількості цих нуклеотидів складаємо пропорцію:
22% - 880
28% - х , звідси х = 1120 Σ (А ) = Σ (Т); ІІ варіант розв'язку
880 - 22% х - 100% х = ( 880 · 100) : 22;
х = 4000;
(А + Т) = 4000 - (Г + Ц) ;
(А + Т) = 4000 - (880 · 2) ;
(А + Т) = 2240 (нуклеотидів) ;
А = Т = 2240 : 2 = 1120 (нуклеотидів) .
2) для визначення довжини ДНК потрібно дізнатися, скільки всього нуклеотидів міститься в 1 ланцюзі:
(880 + 880 + 1120 + 1120) : 2 = 2000 2000 · 0,34 = 680 (нм).
Задача 7. Дана молекула ДНК з відносною молекулярною масою 69 000, з них 8625 припадає на частку аденілових нуклеотидів. Знайдіть кількість всіх нуклеотидів в цій ДНК. Визначте довжину цього фрагмента .
Розв'язок.
1) 69000 : 345 = 200 ( нуклеотидів в ДНК) ,
8625 : 345 = 25 ( аденілових нуклеотидів в цій ДНК) ,
Σ ( Г + Ц) = 200 - ( 25 +25 ) = 150 , тобто їх по 75;
2) 200 нуклеотидів у двох ланцюгах , значить в одній - 100.
100 · 0,34 = 34 (нм).
Задача 8. Ділянка гена має таку послідовність нуклеотидів: ТТТ - ТАЦ - АЦА - ТГТ - ЦАГ. Визначте послідовність нуклеотидів і-РНК і послідовність амінокислот у білковій молекулі, яка синтезується під контролем цього гена .
Розв'язок.
ДНК - ТТТ - ТАЦ - АЦА - ТГТ - ЦАГ і -РНК-ААА - АУГ - УГУ - АЦА - ГУЦ
білок - ліз - мет - цис - тре - вал
Задача 9. Яку довжину має ген, який кодує інсулін, якщо відомо, що молекула інсуліну має 51 амінокислоту, а відстань між нуклеотидами в ДНК складає 0,34 нм?
Розв'язок.
Ген - ділянка ДНК, яка несе інформацію про інсулін. Одну амінокислоту кодує 3 нуклеотиди (триплет) ДНК.
Визначаємо кількість нуклеотидів ДНК, які кодують інсулін:
51· 3 = 153 (нуклеотиди)
Розраховуємо довжину цього гену:
0,34 · 153 = 52,02 (нм).
Задача 10. Дослідження показали, що в і -РНК міститься 34% гуаніну, 18% урацилу, 28% цитозину, 20% аденіну. Визначте відсотковий склад азотистих основ на ділянці ДНК, яка є матрицею для данної і -РНК.
Розв'язок.
Відомо, що 34% гуаніну в і-РНК, тоді в ДНК ланцюзі (кодуючій) буде 34% цитозину, відповідно, 18% урацилу - 18% аденіну, 28% цитозину - 28% гуаніну, 20% аденіну - 20% тиміну (за принципом комплементарності). А + Т та Г + Ц у кодуючій складає:
А + Т = 18% + 20% = 38%, Г + Ц = 28% + 34% = 62%.
У некодуючому ланцюзі ДНК добуткові показники будуть такі ж самі, тільки відсоток окремих основ буде оборотній:
А + Т = 20% + 18% = 38%, Г + Ц = 34% + 28% = 62%.
В обох ланцюгах у парах комплементарних основ буде однакова кількість, тобто А = Т = 19%, Г = Ц = 31%.
Задача 11.
На фрагменті одного ланцюга ДНК нуклеотиди розташовані в послідовності, показаній нижче.
… АГТАЦГГЦАТГТАГЦ …
1. Намалюйте схему структури дволанцюгової молекули ДНК.
2. Якою є довжина в нанометрах цього фрагмента?
3. Якою є маса дволанцюгового фрагмента?
Розв’язання
1. Керуючись властивістю ДНК, здатністю до самовідтворення (реплікації), в основі якого лежить комплементарність, запишемо схему дволанцюгової
ДНК
К А Г Т - А Ц Г - Г Ц А - Т Г Ц - А Г Ц
| | | | | | | | | | | | | | |
Т Ц А - Т Г Ц - Ц Г Т - А Ц Г - Т Ц Г
2. Довжина одного нуклеотида, або відстань між двома сусідніми вздовж осі ДНК, становить 0,34 нм. Довжина дволанцюгового фрагмента дорівнює довжині одного ланцюга.
l = 15 х 0,34 = 5,1 (нм) (15 - кількість нуклеотидів в одному ланцюгу).
3. Середня молекулярна маса одного нуклеотида 345 умовних одиниць, молекулярна маса фрагмента ДНК:
Мr = 345 х 15 = 5175 (а.о.м) (30 – кількість нуклеотидів у двох ланцюгах).
Відповідь. Другий ланцюг фрагмента ДНК має таку структуру: ТЦА – ТГЦ – ЦГТ – АЦГ – ТЦГ ; довжина фрагмента ДНК – 5,1 нм; молекулярна маса фрагмента ДНК – 5175 а.о.м.
Задача 12. У молекулі ДНК 120 гуаніловихнуклеотидів, що становить 15% від загальної кількості. Визначіть кількість інших нуклеотидів, довжину і масу цієї молекули ДНК.
Розв’язання:
1. Г=Ц, тому Г+Ц=120 х 2= 240, або 30%, тоді зможемо знайти загальну кількість нуклеотидів, склавши пропорцію:
240 – 30% x – 100% x =240 x 100/30=800
Знаходимо Т+А=800-240=560, а Т=А=560:2=280.
2. Загальна кількість 800 нуклеотидів, кількість пар 400, тому L(ДНК) = 400 x 0,34=136 нм.
3. M (ДНК) =345x 800=276000 а.о.м .
Відповідь. Загальна кількість нуклеотидів = 800; довжина фрагмента ДНК – 136 нм; молекулярна маса фрагмента ДНК – 276000 а.о.м.
Задача 13.
Біохімічний аналіз показав, що іРНК має 30% аденіну, 18% гуаніну та 20% урацилу. Визначте частку (у %) кожного нуклеотида у відповідному фрагменті дволанцюгової ДНК?
Розв’язання
Дано:
![]() ) =
) =
) : 2 = (30% + 20%) : 2 =
Визначаємо відсоток гуанінового і цитозинових нуклеотидів (окремо) у фрагменті ДНК:
) : 2 = (18% + 32%) : 2 =
кожного нуклеотида у дволанцюгової ДНК
Задача 14.
Фрагмент ланцюга молекули ДНК містить 1100 нуклеотидів , з них 100, 120, і 130 нуклеотидів утворюють інтронні ділянки. Визначте, скільки амінокислот кодує цей фрагмент ДНК:
Розв’язання:
![]() 1). 100 + 120 + 130 = 350 ( кількість N
(інтроннихнуклеотидів) – нуклеотидів, які утворюють інтронні ділянки); 2). 1100
– 350 = 750 ( кількість нуклеотидів, які утворюють екзонні ділянки);
1). 100 + 120 + 130 = 350 ( кількість N
(інтроннихнуклеотидів) – нуклеотидів, які утворюють інтронні ділянки); 2). 1100
– 350 = 750 ( кількість нуклеотидів, які утворюють екзонні ділянки);
3). 750 : 3 = 250 (амінокислотних залишків).
Відповідь. Цей фрагмент ДНК кодує 250 амінокислот.
Задача 15.
Структурний ген (фрагмент молекули ДНК) містить 384 цитозиновихнуклеотидів, що становить 20% від їх загальної кількості. В екзонних ділянках цього гена закодовано білок, який складається із 120 амінокислотних залишків.
1. Який нуклеотидний склад гена?
2. Яка відносна молекулярна маса інтронних ділянок гена?
3. Наскільки зріла іРНК коротша за про-іРНК?
Розв’язання
Дано: 1. Визначаємо загальну кількість
![]() N(Ц-нуклеотидів)
– 384 (20%); нуклеотидів у фрагменті ДНК.
N(амінокислот у білку) – 120; Оскільки на цитозиновінуклеотиди
L(нуклеотида) – 0,34 нм; припадає 20% від їх кількості, то Мr(нуклеотида)
– 345. загальна кількість нуклеотидів становить:
N(Ц-нуклеотидів)
– 384 (20%); нуклеотидів у фрагменті ДНК.
N(амінокислот у білку) – 120; Оскільки на цитозиновінуклеотиди
L(нуклеотида) – 0,34 нм; припадає 20% від їх кількості, то Мr(нуклеотида)
– 345. загальна кількість нуклеотидів становить:
Нуклеотидний склад гена -?
Мr(інтронних ділянок гена) - ? 384 нуклеотидів – 20%; Наскільки зріла іРНК коротша за Х нуклеотидів – 100%;
про-іРНК -? Х
= ![]() =
1920 (нуклеотидів);
=
1920 (нуклеотидів);
За принципом компліментарності:
Г = Ц = 384 нуклеотиди = 20%. Звідси: А = Т = ![]() = 30%.
= 30%.
384 нуклеотиди – 20%;
Х нуклеотидів –
30%; х = ![]() = 576 9нуклеотидів);
= 576 9нуклеотидів);
А = Т 576 нуклеотидів.
2. Знаходимо кількість нуклеотидів у екзонних ділянках гена: 120 3 х 2 = 720 (нуклеотидів).
Знаходимо кількість нуклеотидів у інтронних ділянках гена:
1920 – 720 = 1200 (нуклеотидів).
Знаходимо відносну молекулярну масу інтронних ділянок гена: Mr(інтр. ділянок гена) = 1200 х 345 = 414 000.
3. Довжина молекули про-іРНК дорівнює довжині структурного гена:
l(про-іРНК) = (384 + 576) х 0,34 = 326,4 (нм).
Зріла іРНК складається лише з інформативної частини. Її довжина становить: l(зрілої РНК) = 120 х 3 х 0,34 = 122,4 (нм).
Різниця в довжині про-іРНК та зрілої іРНК складає:
326,4 нм – 122,4 нм = 204 нм.
Відповідь. 1. Ген містить по 576 аденінових і ти мінових нуклеотидів, і по 384 гуанінових і цитозинових нуклеотиди. 2. Відносна молекулярна маса інтронних ділянок гена – 414 000. 3. Різниця в довжині між про- і- РНК та зрілою і- РНК – 204 нм.
Контрольні завдання Задача № 1.
Фрагмент ланцюга А білка нормального гемоглобіну складається із 7 амінокислот, розміщених у такій послідовності:
Вал – лей – лей – тре – про – глн – ліз.
1. Яка будова фрагмента і РНК, що є матрицею для синтезу цього фрагмента молекули гемоглобіну?
2. Яка будова фрагмента ДНК, що кодує дану і РНК ?
Задача № 2.
Фрагмент першого ланцюга ДНК має таку нуклеотидну послідовність: ТАЦАГАТГГАГТЦГЦ. Визначте послідовність мономерів білка, закодованого фрагментом другого ланцюга ДНК.
Задача № 3.
У хворого на синдром Фанконі (порушення утворення кісткової тканини) із сечею виділяються амінокислоти, яким відповідають наступні триплети і-РНК: АУА , ГУЦ, АУГ, УЦА, УУГ, УАУ, ГУУ, АУУ. Визначте, які амінокислоти виділяються із сечею у хворих на синдром Фанконі (див. «Додаток »).
Задача № 4.
У фрагменті ДНК знайдено 1120 аденіновихнуклеотидів, що становить 28% загальної кількості нуклеотидів.
1. Скільки в даному фрагменті міститься гуанінових, цитозинових, ти мінових нуклеотидів?
2. Визначте довжину і відносну молекулярну масу цього фрагмента ДНК.
Задача № 5.
До складу білка входить 800 амінокислот. Визначте довжину гена, який кодує синтез цього білка?
Задача № 6.
Встановлено, що і РНК має 30% аденіну, 18% гуаніну та 20% урацилу. Визначте частку (у %) кожного нуклеотида у відповідному фрагменті дволанцюгової ДНК?
Задача № 7.
Структурний ген (фрагмент молекули ДНК) містить 384 цитозинових нуклеотиди, що становить 20% від їх загальної кількості. В екзонних ділянках цього гена закодовано білок, який складається із 120 амінокислотних залишків.
Задача № 8.
Білок В – мономер. Ген, який кодує цей білок, містить 5 інтронів по 10000 пар нуклеотидів і 4 екзони по 270 пар нуклеотидів кожен.
Задачі з екології
Ланцюг живлення (трофічний ланцюг) — взаємовідносини між організмами під час переносу енергії їжі від її джерела (зеленої рослини) через ряд організмів (шляхом поїдання) на більш високі трофічні рівні.
Ланцюги живлення — це ряди взаємопов’язаних видів, у яких кожний попередній є об’єктом живлення наступного.
Правило екологічної піраміди :кожна з ланок ланцюга живлення може використати лише 5–15 % енергії їжі для побудови речовини свого тіла. Внаслідок неминучої втрати енергії кількість утворюваної органічної речовини в кожній наступній ланці зменшується. Таким чином, кожен ланцюг живлення містить, як правило, не більше ніж 4–5 ланок, оскільки внаслідок втрати енергії загальна біомаса кожної наступної ланки приблизно в 10 разів менша за попередню. Ця закономірність називається правилом екологічної піраміди.
Правило екологічної піраміди:При переході енергії на наступний трофічний рівень лише 10 % від неї використовується для продукування нової біомаси, стаючи запасеною енергією (решта витрачається в процесах метаболізму). Отже, у піраміді
біопродуктивності кожний наступний рівень становить приблизно 10 % від попереднього (100, 10, 1, 0.1, 0.01 % від первинної кількості і т. д.). Екологічна піраміда буває трьох типів:
• піраміда чисел — відображує чисельність окремих організмів на кожному рівні, причому загальне число особин, що беруть участь у ланцюгах живлення, з кожним наступним рівнем зменшується;
• піраміда біомаси — відображує кількісне співвідношення маси органічної речовини на трофічних рівнях; при цьому сумарна маса рослин виявляється більшою, ніж біомаса всіх травоїдних організмів, маса яких, у свою чергу, перевищує масу всіх хижаків;
• піраміда енергії — відображує кількість енергії, яка запасається (в їжі) на кожному рівні, причому на кожному наступному трофічному рівні кількість біомаси, що утворюється за одиницю часу, є більшою, ніж на попередньому. Усі три типи пірамід — продукції, біомаси і чисел — виражають у підсумку енергетичне співвідношення в екосистемах. Перші два правила виявляються в угрупованнях із певною трофічною структурою, останнє (піраміда продукції) має універсальний характер.
Знання законів продуктивності екосистем і кількісний облік потоку енергії мають велике практичне значення. Уміння точно розрахувати потік енергії і масштаби продукції екосистем дозволяє одержати найбільший вихід продукції, необхідної людині.
Приклади розв’язання задач
Задача 1. Біомаса сухого сіна з 1 м2 луки становить 200 г. Використовуючи ланцюг живлення: рослини — корова — людина, розрахуйте скільки гектарів луки необхідно для того, щоб прогодувати людину масою 65 кг (70 % води).
Дано:
m (людини) = 65 кг;
W (H2O) = 70 %;
Трава — корова — людина.
S(гa) -?.
Розв’язок.
1. Розрахуємо суху біомасу кінцевого консумента:
100 % - 70 % = 30 %.
65 кг - 100 %;
Х кг - 30%.
65·30/100 = 19, 5 кг
2. Підставляємо дані в ланцюг живлення і знаходимо біомасу всіх його компонентів:
трава — корова — людина
1950 кг - 195 кг - 19,5 кг
3. Знаходимо площу луки: S= 1950 кг/0,2 кг/м2 = 9750 м2 9750/10000 = 0,975 га.
Відповідь. Щоб прогодувати людину масою 65 кг, потрібен 0,975 га луки.
Задача 2. Користуючись правилом екологічної піраміди визначити, яка площа (в га) біоценозу може прогодувати одну особину останньої ланки в ланцюгу живлення: - планктон – риба - тюлень (300 кг); планктон-нехижі риби-щука (10 кг). Суха біомаса планктону з 1 м2 становить 600 г за рік. Із вказаної у дужках маси 60% становить вода.
Розв’язок.
1. Визначаємо суху масу тюленя: 300 кг - 100%; х кг - 40%: х = 120 кг.
2. На підставі правила екологічної піраміди визначаємо, скільки потрібно планктону: 120 кг – тюлень; 1200 кг – риба; 12000 кг – планктон.
3. Визначаємо площу даного біоценозу, якщо відомо, що суха біомаса планктону з 1 м2 становить 600 г. Отже, 1 м2 - 0,6 кг; х м2 - 1200 кг; х = 20000 м2 = 2 га.
Відповідь: необхідно 2 га планктону.
Задача 3. Біомаса планктону на 1 м2 становить 0,8 кг. Визначити площу (у га) відповідної екосистеми, на якій може прогодуватися пара судаків масою до 0,4 кг кожний, у ланцюзі живлення: планктон – рослиноїдні риби – судак. Із вказаних значень маси риб 40% припадає на суху речовину.
Розв’язання:
1. Визначаємо суху масу тіла 2-х риб:
4кг x 2 = 8 кг
8 кг → 100%
Х кг → 40%
Звідки х = 3,2 кг
2. На підставі правила екологічної піраміди визначаємо масу планктону:
320 кг32 кг3,2 кг
Планктон – рослиноїдні риби – судак
3. Визначаємо площу даного біоценозу, якщо відомо, що суха біомаса планктону з 1 м2 становить 0,8 кг, отже,
1м 2 – 0,8 кг
Х м 2 – 320 кг
Звідки, Х = 400 м2 = 0,04 га.
Відповідь: необхідно 0,04 га водойми.
Контрольні завдання Задача №1.
Яка площа акваторії моря потрібна для прогодування чайки (m = 1 кг, 40% - суха речовина) в ланцюгу живлення: фітопланктон – риба – чайка? Продуктивність фітопланктону – 500 г сухої маси/м 2.
Задача №2.
Яка площа необхідна для існування дельфіна масою 340 кг, з яких 75% - вода, якщо продуктивність біоценозу 1м2 моря 400г сухої біомаси за рік?
Задача №3.
Яка площа здатна прогодувати пару сов масою по 1,5кг кожна, з яких 70% - вода, якщо продуктивність біоценозу 300г сухої біомаси з 1м2 за рік?
Задача №4.
Сарана відклала 80000 яєць, вижило личинок першого віку 24000, другого віку 12000, третього віку 6000, дорослих особин 1200. визначте коефіцієнт виживаності кожної вікової групи і загальний коефіцієнт виживаності.
Задача №5.
Біомаса планктону на 1м 2 становить 400 г. За правилом екологічної піраміди визначте площу (га) відповідного біоценозу, у якому зможе прогодуватися морський леопард масою 300 кг. Ланцюг живлення: планктон – риби – пінгвіни – морський леопард. Із вказаних значень біомаси живих істот 60% припадає на воду.
Задача № 6.
Складіть ланцюг живлення. Користуючись правилом екологічної піраміди, визначте, скільки гектарів луки потрібно, щоб прохарчувати людину масою 60 кг ( із них 66% становить вода ). Суха біомаса трави з 1м2 луки становить 200г за рік.
Задача № 7.
Користуючись правилом екологічної піраміди, визначте, яка площа (в га) відповідного біоценозу може прогодувати одну особину останньої ланки в ланцюзі живлення:
а) планктон – риба – тюлень (300 кг). Суха біомаса планктону з 1м2 моря становить 600 г за рік.
б) планктон – нехижі риби – щука (10 кг). Суха біомаса планктону з 1м2 моря становить 500 г за рік.
в) рослини – заєць – лисиця – вовк (50 кг). Суха біомаса наземної рослинності з 1м2 становить 1000 г за рік. З вказаної у дужках маси 60% становить вода

Додаток 1
Таблиця «Альтернативні ознаки в людини»
|
Домінантна ознака |
Рецесивна ознака |
|
Нормальні |
|
|
Карі очі |
Блакитні очі |
|
Темне волосся |
Світле волосся |
|
Монголоїдний розріз очей |
Європеоїдний розріз очей |
|
Ніс із горбинкою |
Прямий ніс |
|
Широка щілина між різцями |
Вузька щілина між різцями або її відсутність |
|
Зуби великі, виступають вперед |
Звичайна форма і положення зубів |
|
«Ямочки» на щоках |
Відсутність «ямочок» |
|
Біле пасмо волосся |
Рівномірна пігментація волосся |
|
Наявність ластовиння |
Відсутність ластовиння |
|
Мочка вуха вільна |
Мочка вуха приросла |
|
Повні губи |
Тонкі губи |
|
Краще володіння правою рукою |
Краще володіння лівою рукою |
|
Кров резус-позитивна |
Кров резус-негативна |
|
Відчуття смаку фенілтіокарбаміду (ФТК) |
Нездатність відчувати смак ФТК |
|
Патологічні |
|
|
Карликова хондродистрофія |
Нормальний розвиток скелета |
|
Полідактилія (шестипалість) |
Нормальна будова кінцівок |
|
Брахідактилія (короткопалість) |
Нормальна будова кінцівок |
|
Нормальне зсідання крові |
Гемофілія (знижена зсідна здатність крові) |
|
Поліпоз товстої кишки |
Відсутність поліпозу |
|
Нормальне сприймання кольорів |
Дальтонізм (порушення сприймання кольору) |
|
Наявність пігментів |
Альбінізм (відсутність пігментів) |
|
Нормальне засвоєння фенілаланіну |
Фенілкетонурія |
|
Нормальне засвоєння лактози |
Галактоземія |
|
Нормальне засвоєння фруктози |
Фруктозурія |
|
Нормальна будова молекули гемоглобіну |
Серпоподібноклітинна анемія |
Додаток 2
Таблиця генетичного коду
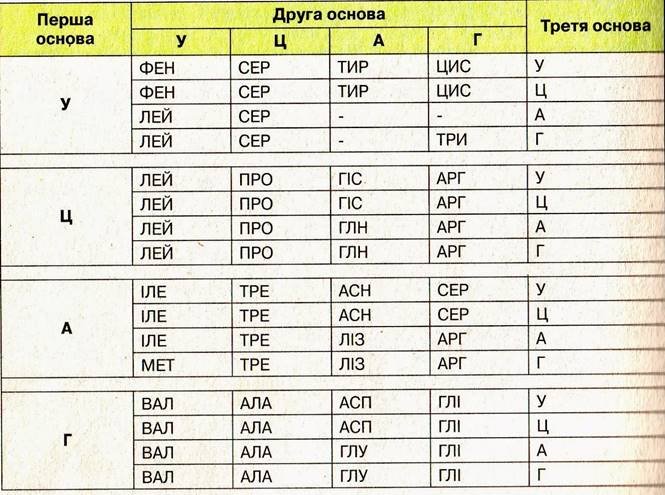
Додаток 3 Назви основних амінокислот та їх позначення
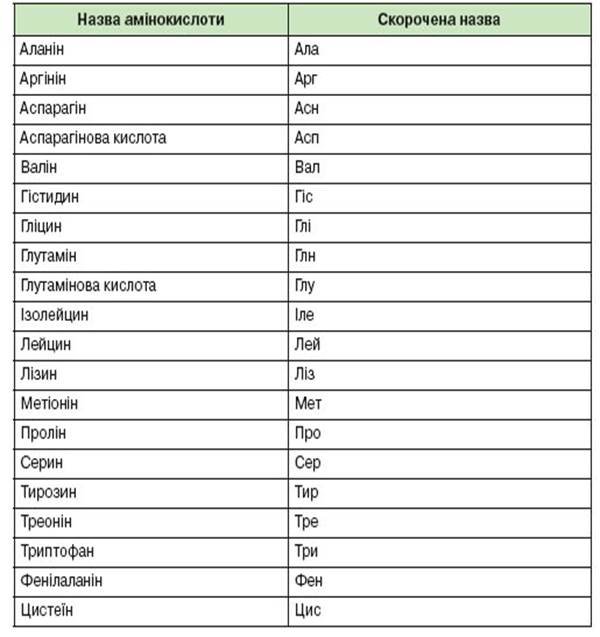
Додаток 4
Гімнастика мозку" - ключ до розвитку здібностей дитини.
Мозок людини є "співдружністю" функціонально асиметричних півкуль - лівого і правого, кожне з яких - не дзеркальне відображення іншого, а необхідне доповнення. Для того, щоб творчо осмислити будь-яку проблему потрібні обидві півкулі: ліва півкуля, логічна, виділяє в кожній проблемі найважливіші, ключові моменти, а праву півкулю, творчу, схоплює її в цілому. Саме права півкуля, завдяки своїй найважливішій функції - інтуїції, допомагає розібратися в ситуації і сформулювати ідею, нехай навіть маячну, але часто нестандартну і нерідко правильну.
Коли півкулі функціонують правильно і між ними зберігається баланс, та взаємодія між ними виражається в ідеальному партнерстві, результатом якого є ефективна творча робота мозку.
Як же забезпечити ефективну взаємодію півкуль і розвинути їх здібності? Ця програма є комплексом вправ,які сприяють розвитку спеціалізації і взаємодії півкуль.
Вправи Гімнастики Мозку спрямовані також на розвиток різних систем координації рухів і психофізичних функцій. "Гімнастика мозку" нагадує звичайну зарядку і проста у виконанні.
Масаж вушних раковин. Помасажуйте мочки вух, потім усю вушну раковину. У кінці вправи розітріть вуха руками.
Колечко. По черзі і якнайшвидше перебирайте пальці рук, сполучаючи в кільце з великим пальцем послідовно вказівний, середній і так далі. Проба виконується в прямому і в зворотному(від мізинця до вказівного пальця) порядку. Спочатку вправа виконується кожною рукою окремо, потім відразу двома руками.
Кулак-ребо-долоня.
Дитині показують три положення руки на площині столу, що послідовно змінюють один одного. Долоня на площині, долоня, стисла в кулак, долоня ребром на площині столу, розпрямлена долоня на площині столу. Дитина виконує пробу разом з педагогом, потім по пам'яті впродовж 8 - 10 повторень моторної програми. Проба виконується спочатку правою рукою, потім - лівою, потім - двома руками разом. При засвоєнні програми або при утрудненнях у виконанні педагог пропонує дитині допомагати собі командами("кулак - ребро - долоня"), вимовними вголос або про себе.
Лезгинка.
Ліву руку складете в кулак, великий палець відставте убік, кулак розгорніть пальцями до себе. Правою рукою прямою долонею в горизонтальному положенні доторкніться до мізинця лівій. Після цього одночасно зміните положення правою і лівою рук. Повторити 6-8 разів. Домагайтеся високої швидкості зміни положень.
Вухо - ніс.
Лівою рукою візьміться за кінчик носа, а правою рукою - за протилежне вухо. Одночасно відпустіть вухо і ніс, хлопніть в долоні, поміняйте положення рук "з точністю до навпаки".
Змійка.
Схрестіть руки долонями один до одного, зчепите пальці в замок. виверніть руки до себе. Рухайте пальцем, який вкаже ведучий. Палець повинен рухатися точно і чітко.Торкатися до пальця не можна. Послідовно у вправі повинні брати участь усі пальці обох рук.
Дзеркальне малювання.
Покладіть на стіл чистий аркуш паперу. Візьміть в обидві руки по олівцю або фломастеру. Почніть малювати одночасно обома руками дзеркально - симетричні малюнки, букви. При виконанні цієї вправи відчуєте, як розслабляються очі і руки. Коли діяльність обох півкуль синхронізується, помітно збільшиться ефективність роботи усього мозку.
"Ледачі вісімки".
На аркуші паперу правою рукою накреслите знак нескінченності(ледачу вісімку), потім те ж саме лівою рукою. А потім правою і лівою одночасно. Малюйте "ледачі вісімки" в повітрі, по черзі кожною рукою і обома одночасно. Ці вправи можна виконувати із студентами під час валеологічних пауз на заняттях.
Список використаних джерел
1. Барна І.В., Барна М. М, Барна Л. С. Біологія: задачі та розв’язки / І.В.Барна. - Тернопіль: Теорія, задачі . розв’язки , 2005. - 382 с.
2. Біологія / За редакцією В.О.Мотузного. - К.: Вища школа, 1991. - 607 с.
3. Ватти К.В., Детье В.Н. Биология (биологические процессьі и законьї) / К.В.Ватти, В.Н.Детье. - М.: Мир, 1975. - 822 с.
4. Данилова О.В. Біологія: зб.тест. завдань для підготовки до зовн. незалеж. оцінювання. - 2 - ге вид., доповнене / О.В.Данилова, С.А.Данилов, В.П.Дмитренко. - К.: Генеза, 2011. - 128 с.
5. Довідник з біології/ За редакцією академіка НАН України К.М.Ситника. - К.: Наукова думка, 1998. - 682 с.
6. Загальна біологія: [Підручн. для учнів 10 - 11- х кл. серед, загальноосвітніх шкіл] / М.Є.Кучеренко, Ю.Г.Вервес, П.Г.Балан та ін. - К.: Генеза, 2000. - 464 с.
7. Овчинніков С.О. Збірник задач і вправ із загальної біології / С.О.Овчинніков. - К.: Генеза, 2000. - 150 с.
8. Прокопенко Л.І. Біологія: Довідник для абітурієнтів та школярів загальноосвітніх шкіл К: Генеза , 2000.-150 с.
9. Барна І.В. Загальна біологія: збірник задач / І.В.Барна. - Тернопіль: Підручники і посібники, 2006. - 736 с.


про публікацію авторської розробки
Додати розробку
-

Мітіна Світлана Миколаївна
05.01.2021 в 12:12
Загальна:
5.0
Структурованість
5.0
Оригінальність викладу
5.0
Відповідність темі
5.0
-

Крапивна Алла Володимирівна
11.10.2020 в 19:41
Загальна:
4.0
Структурованість
5.0
Оригінальність викладу
3.0
Відповідність темі
4.0
-

Мігунова Олена Ігорівна
20.02.2019 в 07:55
Загальна:
4.0
Структурованість
4.0
Оригінальність викладу
4.0
Відповідність темі
4.0
-

Стефанова Світлана Володимирівна
09.10.2018 в 21:38
-
Загальна:
4.3
Структурованість
4.0
Оригінальність викладу
4.0
Відповідність темі
5.0
-

Шишковська Людмила Григорівна
04.10.2018 в 16:20
Чудово.
Загальна:
4.0
Структурованість
4.0
Оригінальність викладу
4.0
Відповідність темі
4.0
-

Слівінська Ольга Лонгинівна
01.10.2018 в 23:49
Спасибі
Загальна:
4.3
Структурованість
4.0
Оригінальність викладу
4.0
Відповідність темі
5.0
Показати ще 3 відгука