Презентація "Загальний план будови клітин"


Вчитель: Теслюк Н.Ю., ЗЗСО Соснівський ліцей №3

ОСВІТНІ ЦІЛІ: розвинути поняття про клітинний рівень життя; ознайомити учнів з етапами розвитку цитології та методами цитологічних досліджень; сформулювати основні положення клітинної теорії; розвинути знання учнів про будову клітин прокаріот і еукаріот; сформувати практичні вміння самостійно досліджувати ці клітини і вивчити їхню будову; порівнювати і робити висновки про еволюцію клітин; на прикладі самовідданої праці вчених виховувати учнів.

Кліти́на (від лат. cellula — комірка) — структурно-функціональна одиниця всіх живих організмів, для якої характерний власний метаболізм та здатність до відтворення. Від середовища, яке її оточує, клітина відмежована плазматичною мембраною (плазмалемою). Розрізняють два основні типи клітин: прокаріотичні, що не мають сформованого ядра, характерні для бактерій та архей, та еукаріотичні, в яких наявне ядро, властиві для всіх інших клітинних форм життя, зокрема рослин, грибів та тварин. До неклітинних форм життя належать лише віруси, але вони не мають власного метаболізму і не можуть розмножуватись поза межами клітин-господарів. Усі організми поділяються на одноклітинні, колоніальні та багатоклітинні. До одноклітинних належать бактерії, археї, деякі водорості і гриби, а також найпростіші. Колоніальні та багатоклітинні організми складаються з великої кількості клітин. Різниця між ними полягає в тому, що колоніальні організми складаються з недиференційованих або слабо диференційованих клітин, які можуть виживати одна без одної. Клітини багатоклітинних організмів більш-менш спеціалізовані на виконанні певних функцій і залежні одна від одної в процесах життєдіяльності. До багатоклітинних організмів належить зокрема і людина, тіло якої складається приблизно з 1013 клітин.

ІСТОРІЯ ВІДКРИТТЯ ТА ДОСЛІДЖЕННЯ КЛІТИН Більшість еукаріотичних клітин мають розміри до 100 мкм, а прокаріотичні ще на порядок менші, тому людина не може бачити їх неозброєним оком. Відкриття та дослідження клітин стало можливим тільки після винайдення Янсеном оптичного мікроскопа (1590 року). До найважливіших подій, пов'язаних із раннім розвитком клітинної біології належать: 1665 — Роберт Гук вперше побачив мертві клітини, вивчаючи будову корка під мікроскопом. Гук вважав, що клітини порожні, а живою речовиною є клітинні стінки. 1650—1700 — Антоні ван Левенгук вперше спостерігав під мікроскопом живі клітини, зокрема найпростіші, а також еритроцити. 1831—1839 — Роберт Браун описав ядро, як сферичне тільце, наявне в рослинних клітинах. 1838—1839 — ботанік Матіас Шлейден і зоолог Теодор Шванн, об'єднавши ідеї різних вчених, створили клітинну теорію, згідно з якою клітина є основною структурною та функціональною одиницею живих організмів. 1840 — Пуркіньє запропонував назву протоплазма для позначення клітинного вмісту, переконавшись у тому, що саме вміст, а не клітинні стінки, є живою речовиною. 1855 — Вірхов довів, що всі клітини утворюються із інших клітин шляхом поділу. 1866 — Геккель встановив, що збереження та передачу спадкових ознак здійснює ядро. 1866—1898 — описано основні компоненти клітини, які можна побачити під оптичним мікроскопом. Цитологія набуває характеру експериментальної науки. 1900 — за появою генетики починає розвиватись цитогенетика, що вивчає поведінку хромосом під час поділу та запліднення, її вплив на спадкові ознаки організмів. 1946 — у біології розпочалося використання електронного мікроскопа, що дозволило вивчати ультраструктуру клітин.

Рисунок клітин корка із праці «Мікрографія» Роберта Гука

КЛІТИННА ТЕОРІЯ Клітинну теорію сформульовано у 1838—1839 роках ботаніком Матіасом Шлейденом і зоологом Теодором Шванном. Ці вчені довели принципову подібність між собою тваринних та рослинних клітин, і на основі всіх накопичених до того часу знань постулювали, що клітина є структурною та функціональною одиницею всіх живих організмів. 1855 року Рудольф Вірхов доповнив клітинну теорію твердженням лат. «Omnis cellula eх cellula» — «Кожна клітина — з клітини». Клітинна теорія є однією із основоположних ідей сучасної біології, вона стала незаперечним доказом єдності всього живого та фундаментом для розвитку таких дисциплін як ембріологія, гістологія та фізіологія. Основні положення клітинної теорії не втратили своєї актуальності, проте від часу створення її було доповнено, і наразі вона містить такі твердження: Клітина — елементарна одиниця будови, функціонування, розмноження і розвитку всіх живих організмів, поза межами клітини немає життя. Клітина — цілісна система, що містить велику кількість пов'язаних один з одним елементів — органел. Клітини різних організмів схожі (гомологічні) за будовою та основними властивостями і мають спільне походження. Збільшення кількості клітин відбувається шляхом їх поділу, після реплікації її ДНК: клітина — від клітини. Багатоклітинний організм — це нова система, складний ансамбль із великої кількості клітин, об'єднаних та інтегрованих у системи тканин і органів, пов'язаних між собою за допомогою хімічних факторів: гуморальних і нервових. Клітини багатоклітинних організмів мають однаковий набір генетичної інформації, але відрізняються за рівнем експресії (роботи) окремих генів, що призводить до їх морфологічної та функціональної різноманітності — диференціації. Слід зазначити, що в різних джерелах кількість та формулювання окремих положень сучасної клітинної теорії можуть відрізнятись.

МЕТОДИ ДОСЛІДЖЕННЯ КЛІТИН Вперше клітини вдалось побачити тільки після створення світлових мікроскопів, з того часу і до цих пір мікроскопія залишається одним із найважливіших методів дослідження клітин. Використовується світлова (оптична) мікроскопія, що попри свою порівняно невелику роздільну здатність має ту перевагу, що дозволяє спостерігати за живими клітинами. У ХХ столітті була винайдена електронна мікроскопія, що дала можливість вивчити ультраструктуру клітин. Для вивчення функцій клітин та їх частин використовують різноманітні біохімічні методи як препаративні, наприклад фракціонування методом диференційного центрифугування, так і аналітичні. Для експериментальних та практичних цілей використовують методи клітинної інженерії. Всі згадані методичні підходи можуть використовуватись у поєднанні із методами культури клітин. Копія мікроскопа, що застосовувався Левенгуком

ОПТИЧНА МІКРОСКОПІЯ У оптичному мікроскопі збільшення об'єкта досягається завдяки серії лінз, через які проходить світло. Максимальне збільшення, яке можна досягнути завдяки оптичному мікроскопу становить близько 1000. Ще однією важливою характеристикою є роздільна здатність — відстань між двома точками, які ще розпізнаються окремо, іншими словами роздільна здатність характеризує чіткість зображення. Ця величина обмежується довжиною світлової хвилі, навіть при використанні найбільш короткохвильового світла — ультрафіолетового — можна досягнути тільки роздільної здатності близько 200 нм, таку роздільність було отримано ще в кінці XIX століття. Таким чином найменші структури, які можна спостерігати під оптичним мікроскопом це мітохондрії і невеликі бактерії, лінійний розмір яких становить приблизно 500 нм. Проте об'єкти, менші за 200 нм, видні у світловому мікроскопі, якщо вони самі випромінюють світло. Ця особливість використовується у флуоресцентній мікроскопії, за якої клітинні структури чи окремі білки зв'язуються зі спеціальними флуоресцентними білками або антитілами із флуоресцентними мітками. На якість зображення, отриманого за допомогою оптичного мікроскопа, впливає також контрастність, її можна збільшити використовуючи різні методи забарвлення клітин. Для вивчення живих клітин використовують фазовоконтрастну та диференційну інтерференційно-кантрастну і темнопольну мікроскопію. Конфокальні мікроскопи дозволяють покращити якість флуоресцентних зображень.
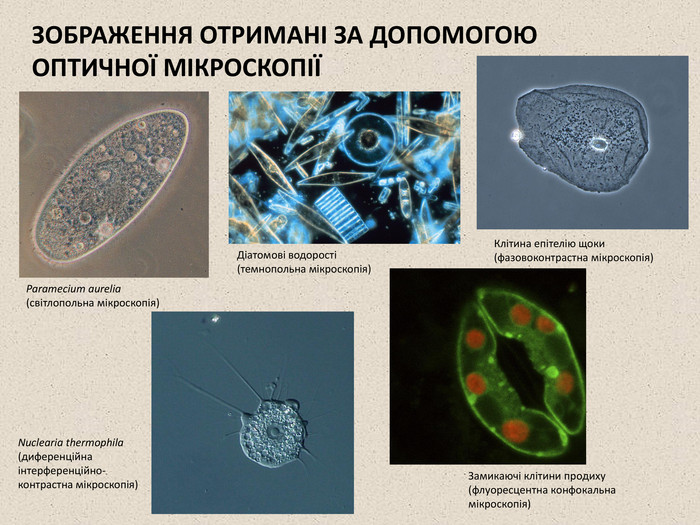
ЗОБРАЖЕННЯ ОТРИМАНІ ЗА ДОПОМОГОЮ ОПТИЧНОЇ МІКРОСКОПІЇ Paramecium aurelia (світлопольна мікроскопія) Діатомові водорості (темнопольна мікроскопія) Клітина епітелію щоки (фазовоконтрастна мікроскопія) Nuclearia thermophila (диференційна інтерференційно- контрастна мікроскопія) Замикаючі клітини продиху (флуоресцентна конфокальна мікроскопія)

ЕЛЕКТРОННА МІКРОСКОПІЯ У 30-их роках XX століття був сконструйований електронний мікроскоп, в якому замість світла через об'єкт пропускається пучок електронів. Теоретична межа роздільності для сучасних електронних мікроскопів становить близько 0,002 нм, проте із практичних причин для біологічних об'єктів досягається роздільність тільки близько 2 нм. За допомогою електронного мікроскопа можливо вивчати ультраструктуру клітин. Розрізняють два основні типи електронної мікроскопії: скануючу та трансмісійну. Скануюча електронна мікроскопія (СЕМ) використовується для вивчення поверхні об'єкта. Зразки найчастіше покривають тонкою плівкою золота. СЕМ дозволяє отримувати об'ємні зображення. Трансмісійна електронна мікроскопія (ТЕМ) — використовується для вивчення внутрішньої будови клітини. Пучок електронів пропускається через об'єкт, що попередньо обробляється важкими металами, які накопичуються у певних структурах збільшуючи їхню електронну густину. Електрони розсіюються на ділянках клітини з більшою електронною густиною, внаслідок чого на зображеннях ці області виглядають темнішими
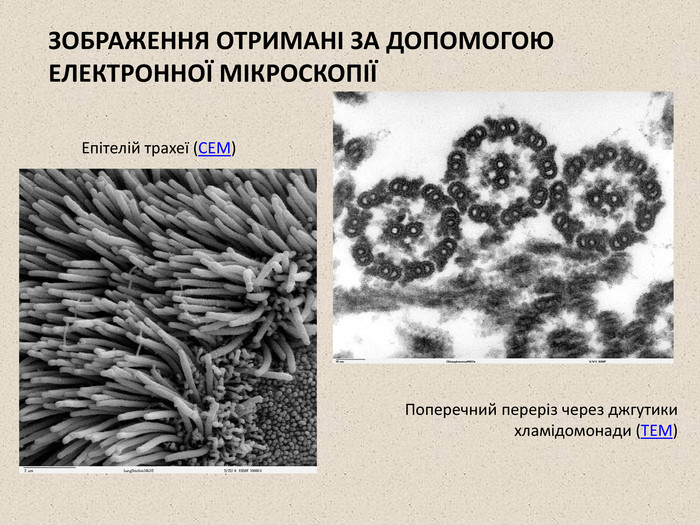
ЗОБРАЖЕННЯ ОТРИМАНІ ЗА ДОПОМОГОЮ ЕЛЕКТРОННОЇ МІКРОСКОПІЇ Епітелій трахеї (СЕМ) Поперечний переріз через джгутики хламідомонади (ТЕМ)

ФРАКЦІОНУВАННЯ КЛІТИН Для встановлення функцій окремих компонентів клітини важливо виділити їх у чистому вигляді, найчастіше це робиться за допомогою методу диференційного центрифугування. Розроблені методики, що дозволяють отримати чисті фракції будь-яких клітинних органел. Отримання фракцій починається із руйнування плазмалеми і утворення гомогенату клітин. Гомогенат послідовно центрифугується при різних швидкостях, на першому етапі можна отримати чотири фракції: (1) ядер і великих уламків клітин, (2) мітохондрій, пластид, лізосом і пероксисом, (3) міркосом — пухирців апарату Гольджі та ендоплазматичного ретикулуму, (4) рибосом, в супернатанті залишаться білки та дрібніші молекули. Подальше диференційне центрифугування кожної із змішаних фракцій дозволяє отримати чисті препарати органел, до яких можна застосовувати різноманітні біохімічні та мікроскопічні методи
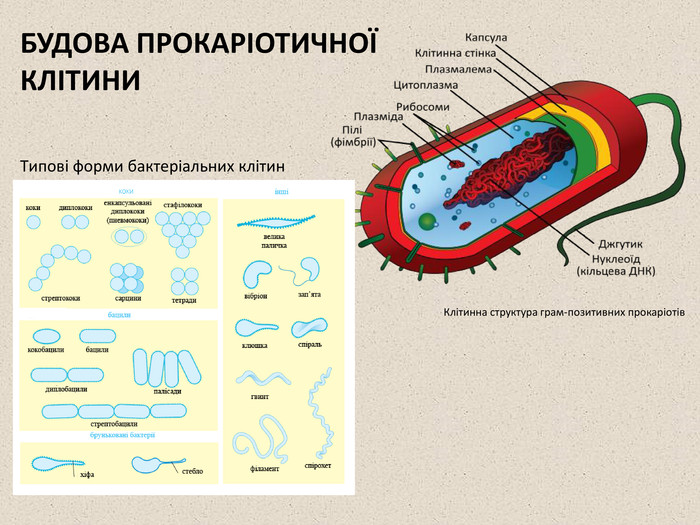
БУДОВА ПРОКАРІОТИЧНОЇ КЛІТИНИ Типові форми бактеріальних клітин Клітинна структура грам-позитивних прокаріотів

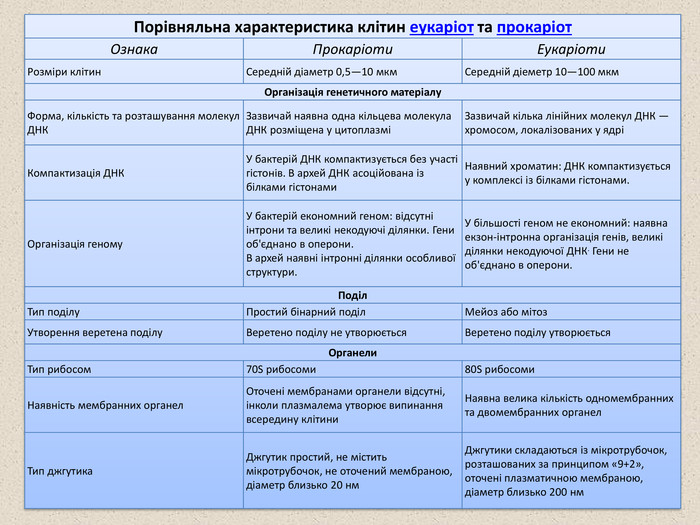
Клітини двох основних груп прокаріот: бактерій та архей схожі за структурою, найхарактернішими їх ознаками є відсутність ядра та мембранних органел. Основними компонентами прокаріотичної клітини є: 1. Клітинна стінка оточує клітинну ззовні, захищає її, надає сталої форми, запобігає осмотичному руйнуванню. У бактерій клітинна стінка складається із пептидоглікану (муреїну), що побудований із довгих полісахаридних ланцюгів, з'єднаних між собою короткими пептидними перемичками. За будовою клітинної стінки розрізняють дві групи бактерій: Грам-позитивні бактерії (наприклад роди Staphylococcus, Bacillus, Lactobacillus) мають простішу структуру клітинної сінки, що складається майже виключно із муреїну; У грам-негативних бактерій (наприклад роди Salmonella, Escherichia, Azotobacter) клітинна стінка містить менше пептидоглікану, і має додаткову зовнішню мембрану, що складається із фосфоліпідів. Клітинна стінка архей не містить муреїну, а побудована, здебільшого, з різноманітних білків та полісахаридів. 2. Капсула — наявна у деяких бактерій слизова оболонка, розташована зовні від клітинної стінки. Складається, здебільшого, з різноманітних білків, вуглеводів та уронових кислот. Капсули захищають клітини від висихання, можуть допомагати бактеріям у колоніях утримуватись разом, а індивідуальним бактеріям — прикріплюватись до різних субстратів. Окрім цього капсули надають клітині додатковий захист: наприклад капсульовані штами пневмококів вільно розмножуються в організмі та викликають запалення легень, тоді як не капсульовані швидко знищуються імунною системою та є абсолютно нешкідливими. Структура клітинної стінки двох типів бактерій
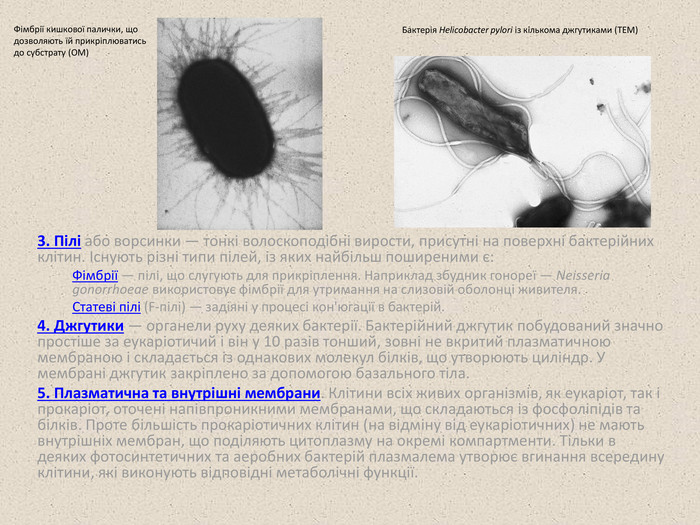
3. Пілі або ворсинки — тонкі волоскоподібні вирости, присутні на поверхні бактерійних клітин. Існують різні типи пілей, із яких найбільш поширеними є: Фімбрії — пілі, що слугують для прикріплення. Наприклад збудник гонореї — Neisseria gonorrhoeae використовує фімбрії для утримання на слизовій оболонці живителя. Cтатеві пілі (F-пілі) — задіяні у процесі кон'югації в бактерій. 4. Джгутики — органели руху деяких бактерії. Бактерійний джгутик побудований значно простіше за еукаріотичий і він у 10 разів тонший, зовні не вкритий плазматичною мембраною і складається із однакових молекул білків, що утворюють циліндр. У мембрані джгутик закріплено за допомогою базального тіла. 5. Плазматична та внутрішні мембрани. Клітини всіх живих організмів, як еукаріот, так і прокаріот, оточені напівпроникними мембранами, що складаються із фосфоліпідів та білків. Проте більшість прокаріотичних клітин (на відміну від еукаріотичних) не мають внутрішніх мембран, що поділяють цитоплазму на окремі компартменти. Тільки в деяких фотосинтетичних та аеробних бактерій плазмалема утворює вгинання всередину клітини, які виконують відповідні метаболічні функції. Фімбрії кишкової палички, що дозволяють їй прикріплюватись до субстрату (ОМ) Бактерія Helicobacter pylori із кількома джгутиками (ТЕМ)

6. Нуклеоїд — не відмежована мембранами ділянка цитоплазми, в якій розташовано кільцеву молекулу ДНК — «бактерійну хромосому», де зберігається весь генетичний матеріал клітини. 7. Плазміди — невеликі додаткові кільцеві молекули ДНК, що несуть зазвичай всього декілька генів. Плазміди, на відміну від бактерійної хромосоми, не є обов'язковим компонентом клітини. Зазвичай вони надають бактерії певних корисних для неї властивостей, таких як стійкість до антибіотиків, здатність засвоювати з середовища певні енергетичні субстрати, здатність ініціювати статевий процес тощо. 8. Рибосоми прокаріот, як і в усіх інших живих організмів, відповідають за здійснення процесу трансляції (одного із етапів біосинтезу білка). Проте бактерійні хромосоми дещо менші за еукаріотичні (коефіцієнти седиментації 70S та 80S відповідно) і мають інший склад білків та РНК. Через це бактерії, на відміну від еукаріот, чутливі до таких антибіотиків як еритроміцин та тетрациклін, що вибірково діють на 70S рибосоми. 9. Ендоспори — оточені щільною оболонкою структури, що містять ДНК бактерії і забезпечують виживання у несприятливих умовах. До утворення ендоспор здатні лише деякі види прокаріот, наприклад представники родів Clostridium (C. Tetani — збудник правцю, C. botulinum — збудник ботулізму, C. Perfringens — збудник газової гангрени тощо) та Bacillus (зокрема B. anthracis — збудник сибірської виразки). Для утворення ендоспори клітина реплікує свою ДНК і оточує копію щільною оболонкою, з утвореної структури видаляється надлишок води, і в ній сповільнюється метаболізм. Спори бактерій можуть витримувати досить жорсткі умови середовища, такі як тривале висушування, кип'ятіння, короткохвильове опромінення тощо Збудник правцю — Clostridium tetani, у клітинах видно ендоспори (оранжеві)

БУДОВА ЕУКАРІОТИЧНОЇ КЛІТИНИ До еукаріот належать три основні царства живих організмів: Тварини, Рослини та Гриби. Попри деякі відмінності у будові, їхні клітини схожі між собою і відрізняються від клітин прокаріот наявністю ядра та компартменталізацією цитоплазми на окремі відсіки за допомогою системи внутрішніх мембран. Живий вміст клітини називається протоплазмою, протоплазму оточено напівпроникною плазматичною мембраною або плазмалемою, зовні протоплазми можуть розташовуватись надмембранні структури, такі як клітинна стінка (у рослин та грибів) або глікокалікс (у тварин). До складу протоплазми клітини входить ядро та цитоплазма, яка у свою чергу складається із колоїдного розчину — гіалоплазми — та розміщених у ній органел — постійних структурних і функціональних елементів клітини. Окрім цього клітини можуть тимчасово накопичувати певні речовини, що утворюють клітинні включення.
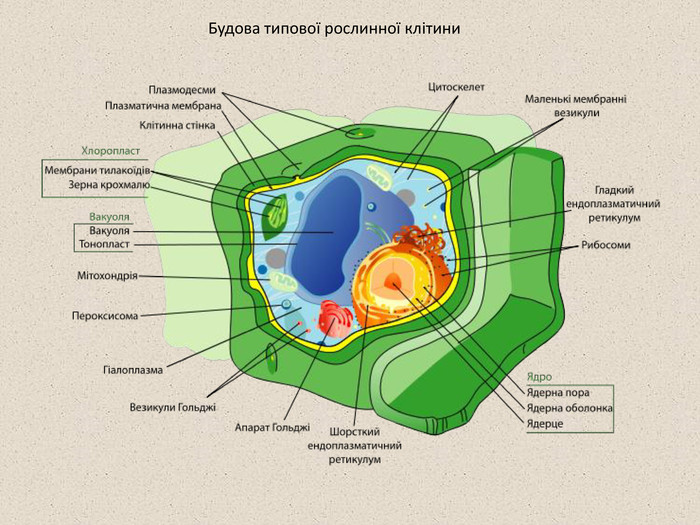
Будова типової рослинної клітини

Будова типової тваринної клітини
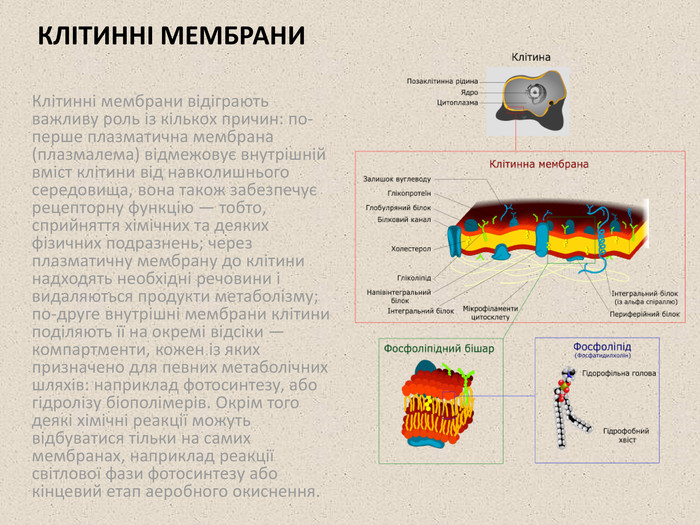
КЛІТИННІ МЕМБРАНИ Клітинні мембрани відіграють важливу роль із кількох причин: по-перше плазматична мембрана (плазмалема) відмежовує внутрішній вміст клітини від навколишнього середовища, вона також забезпечує рецепторну функцію — тобто, сприйняття хімічних та деяких фізичних подразнень; через плазматичну мембрану до клітини надходять необхідні речовини і видаляються продукти метаболізму; по-друге внутрішні мембрани клітини поділяють її на окремі відсіки — компартменти, кожен із яких призначено для певних метаболічних шляхів: наприклад фотосинтезу, або гідролізу біополімерів. Окрім того деякі хімічні реакції можуть відбуватися тільки на самих мембранах, наприклад реакції світлової фази фотосинтезу або кінцевий етап аеробного окиснення.

Товщина біологічних мембран становить близько 7 нм. Вони складаються в основному із фосфоліпідів, у молекулах яких виділяють дві основні частини: гідрофільну «голову» (залишок фосфатної кислоти і холіну, серину, етаноламіну або іншої полярної сполуки) та два гідрофобні «хвости» (залишки жирних кислот). У складі клітинної мембрани фосфоліпіди утворюють бішар, в якому гідрофільні голови повернуто назовні — у полярний водний розчин, а гідрофобні хвости — всередину. Крім фосфоліпідів до складу мембран входять також інші типи ліпідів, такі як гліколіпіди, сфінголіпіди та холестерол. Іншим важливим компонентом клітинної мембрани є білки, їх вміст може коливатись від 18% (у мембрані аксона) до 75% (у мембранах тилакоїдів). Усі мембранні білки можна поділити на три основні групи: Інтегральні мембранні білки мають у своєму складі гідрофобний домен, який повністю або частково пронизує ліпідний бішар, до цього класу належать усі іонні канали та більшість клітинних рецепторів; Білки, заякорені в мембрані ковалентно, з'єднані із певною неполярною сполукою, найчастіше — глікозилфосфатидилінозитолом (англ. GPI), яка входить у гідрофобну частину ліпідного бішару, і таким чином утримує білок на поверхні мембрани. Представниками цієї групи є G-білки; Периферійні білки нековалентно взаємодіють з іншими білками або гідрофільними головами фосфоліпідів і утримуються поблизу поверхні мембрани тільки тимчасово. Прикладом цієї групи білків можуть бути деякі ферменти. 1972 року Сінгер і Ніколсон запропонували рідинно-мозаїчну модель будови мембрани, згідно із якою фосфоліпідний бішар є двовимірною рідиною, в якій вільно плавають білкові молекули, утворюючи ніби мозаїку, яка постійно змінюється. Зовнішня і внутрішня сторони мембрани відрізняються фосфоліпідним і білковим складом та функціями. Схема перерізу ліпідного бішару перпендикулярно площині мембрани

До основних функцій мембран належать: 1. Обмеження вмісту клітини. Мембрани характеризуються вибірковою проникністю: через них можуть проходити неполярні речовини (наприклад кисень, вуглекислий газ, стероїдні гормони), але не великі полярні та заряджені молекули (амінокислот, моносахаридів, неорганічних іонів). Маленькі полярні молекули, такі як вода, здатні перетинати ліпідний бішар, але цей процес ускладнено. Через такі властивості мембрана утримує всередині клітини всі біополімери та заряджені молекули, а також запобігає потраплянню таких молекул зовні. 2. Транспорт. Мембрани регулюють процес транспорту до клітини потрібних речовин та виведення із неї відходів. Розрізняють наступні види клітинного транспорту: Пасивний транспорт — рух речовин через мембрану за градієнтом концентрації (від ділянки з більшою концентрацією до ділянок із меншою концентрацією) без витрат енергії: Проста дифузія — рух речовин (наприклад гідрофобних молекул або розчинених газів) безпосередньо через ліпідний бішар. Різновидом простої дифузії є осмос — рух води через напівпроникну мембрану з ділянки з меншою концентрацією розчину до ділянки з більшою концентрацією. Полегшена дифузія — рух молекул через особливі білкові канали або за посередництва білкових переносників за градієнтом концентрації (наприклад рух неорганічних іонів через іонні канали). Активний транспорт — рух речовин через мембрану проти градієнту концентрації, що відбувається із витратою енергії та здійснюється за допомогою спеціальних білків-насосів. Розрізняють первинний активний транспорт, для якого використовується енергія гідролізу АТФ (наприклад робота натрій-калієвого насосу), та вторинний активний транспорт, за якого для транспорту однієї речовини проти градієнту концентрації інша транспортується за градієнтом (наприклад процес всмоктування глюкози клітинами тонкого кишківника). Ендоцитоз та екзоцитоз — енерговитратні процеси транспортування речовин і часточок у клітину (ендоцитоз) або з клітини (екзоцитоз) за участі мембранних везикул (пухирців). 3. Рецепція. На поверхні плазматичної мембрани розташовано велику кількість клітинних рецепторів (найчастіше глікопротеїнів), що сприймають різні хімічні та фізичні сигнали та передають їх всередину клітини. Таким чином клітина сприймає сигнали, що передаються у формі гормонів, нейромедіаторів, цитокінів. За участі рецепторів та інших поверхневих білків клітини багатоклітинного організму розпізнають одна одну. 4. Метаболічна функція. Багато з мембранних білків є ферментами, інколи кілька мембранозв'язаних ферментів можуть бути організовані у комплекси для здійснення послідовних метаболічних реакцій, при цьому мембрана виступає каркасом для їх просторової організації. Реакції світлової фази фотосинтезу та електронтранспортного ланцюга мітохондрій можуть відбуватись тільки на відповідних мембранах. 5. Клітинна адгезія. Мембрани тварин, зокрема деякі мембранні білки, такі як кадгерини, забезпечують прикріплення клітин багатоклітинного організму одна до одної або до позаклітинного матриксу. Таким чином забезпечується структурна цілісність тканин тваринного організму. Контакт із мікрооточенням за участі мембранних білків також є важливим для виживання багатьох типів клітин, без нього вони гинуть шляхом апоптозу.
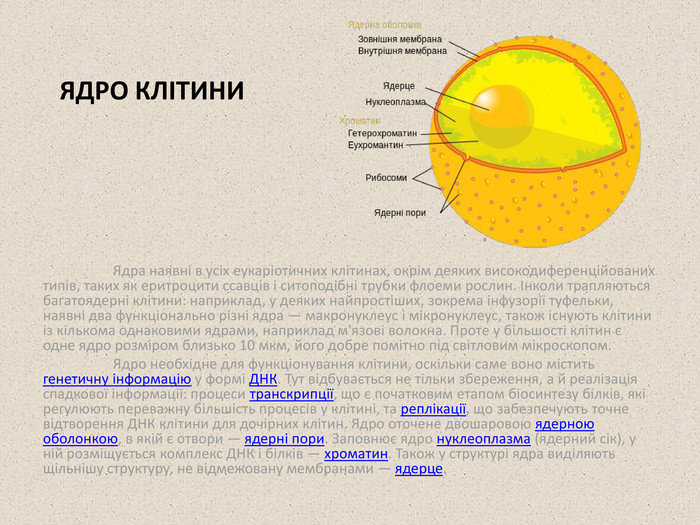
ЯДРО КЛІТИНИ Ядра наявні в усіх еукаріотичних клітинах, окрім деяких високодиференційованих типів, таких як еритроцити ссавців і ситоподібні трубки флоеми рослин. Інколи трапляються багатоядерні клітини: наприклад, у деяких найпростіших, зокрема інфузорії туфельки, наявні два функціонально різні ядра — макронуклеус і мікронуклеус, також існують клітини із кількома однаковими ядрами, наприклад м'язові волокна. Проте у більшості клітин є одне ядро розміром близько 10 мкм, його добре помітно під світловим мікроскопом. Ядро необхідне для функціонування клітини, оскільки саме воно містить генетичну інформацію у формі ДНК. Тут відбувається не тільки збереження, а й реалізація спадкової інформації: процеси транскрипції, що є початковим етапом біосинтезу білків, які регулюють переважну більшість процесів у клітині, та реплікації, що забезпечують точне відтворення ДНК клітини для дочірних клітин. Ядро оточене двошаровою ядерною оболонкою, в якій є отвори — ядерні пори. Заповнює ядро нуклеоплазма (ядерний сік), у ній розміщується комплекс ДНК і білків — хроматин. Також у структурі ядра виділяють щільнішу структуру, не відмежовану мембранами — ядерце.

Ядерна оболонка та ядерні пори Ядерна оболонка складається з двох мембран: зовнішня безпосередньо переходить в ендоплазматичний ретикулум і може бути всіяна рибосомами; внутрішня має спеціальні білки, до яких приєднуються філаменти ядерної пластинки (ламіни) — структури, що підтримує форму ядра. Між зовнішньою та внутрішньою мембранами розташований перинуклеарний простір неперервний із внутрішнім простором ендоплазматичного ретикулуму. У деяких місцях зовнішня та внутрішня мембрани ядра зливаються, утворюючи отвори діаметром близько 100 нм, ці отвори називаються ядерними порами. Всередині кожної пори розміщено складний апарат із молекул близько 30 різних білків нуклеопоринів — ядерний поровий комплекс, що регулює транспорт між ядром і цитоплазмою. За секунду ядерна пора може переносити більше 500 макромолекул у двох напрямках одночасно. До ядра транспортуються переважно білки — гістони, рибосомальні білки, ферменти, що беруть участь в процесах транскрипції, реплікації, репарації, регуляторні молекули а також різні метаболіти, такі як нуклеотиди. Із ядра до цитоплазми транспортуються зрілі молекули мРНК, субодиниці рибосом. Під час клітинного поділу ядерна оболонка зникає.
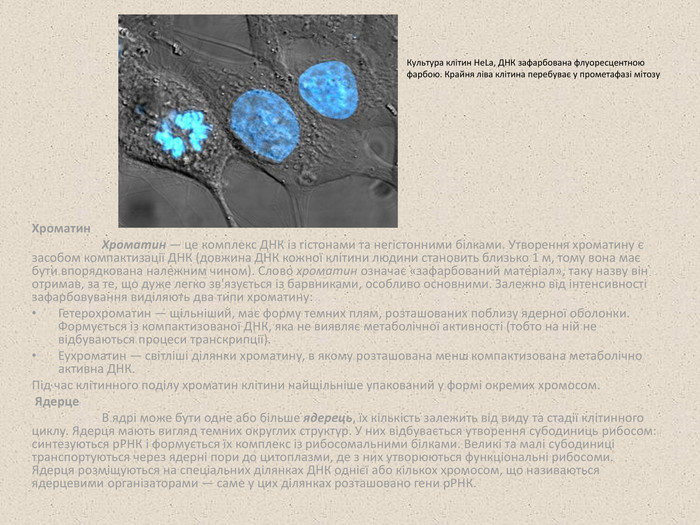
Хроматин Хроматин — це комплекс ДНК із гістонами та негістонними білками. Утворення хроматину є засобом компактизації ДНК (довжина ДНК кожної клітини людини становить близько 1 м, тому вона має бути впорядкована належним чином). Слово хроматин означає «зафарбований матеріал», таку назву він отримав, за те, що дуже легко зв'язується із барвниками, особливо основними. Залежно від інтенсивності зафарбовування виділяють два типи хроматину: Гетерохроматин — щільніший, має форму темних плям, розташованих поблизу ядерної оболонки. Формується із компактизованої ДНК, яка не виявляє метаболічної активності (тобто на ній не відбуваються процеси транскрипції). Еухроматин — світліші ділянки хроматину, в якому розташована менш компактизована метаболічно активна ДНК. Під час клітинного поділу хроматин клітини найщільніше упакований у формі окремих хромосом. Ядерце В ядрі може бути одне або більше ядерець, їх кількість залежить від виду та стадії клітинного циклу. Ядерця мають вигляд темних округлих структур. У них відбувається утворення субодиниць рибосом: синтезуються рРНК і формується їх комплекс із рибосомальними білками. Великі та малі субодиниці транспортуються через ядерні пори до цитоплазми, де з них утворюються функціональні рибосоми. Ядерця розміщуються на спеціальних ділянках ДНК однієї або кількох хромосом, що називаються ядерцевими організаторами — саме у цих ділянках розташовано гени рРНК. Культура клітин HeLa, ДНК зафарбована флуоресцентною фарбою. Крайня ліва клітина перебуває у прометафазі мітозу

ЦИТОПЛАЗМА КЛІТИНИ Цитоплазма клітини складається із водянистої основної речовини — гіалоплазми, в якій розташовано органели, нитки цитоскелету та (інколи) клітинні включення. Гіалоплазма або основна речовина цитоплазми приблизно на 90% складається з води, в якій розчинено всі основні біомолекули: солі, цукри, амінокислоти, нуклеотиди, вітаміни та гази утворюють істинний розчин, тоді як великі молекули, зокрема білки, перебувають у колоїдному розчині. У гіалоплазмі відбувається велика кількість метаболічних процесів, зокрема гліколіз. Вона може змінювати свої властивості, переходячи зі стану золю до стану густішого гелю. Спостерігаючи за живою цитоплазмою клітини, зазвичай, можна помітити, що вона рухається, найкраще видно рух мітохондрій і пластид, це явище називають циклозом.

РИБОСОМИ Рибосоми — немембранні органели клітини, що складаються з рРНК та рибосомних білків (протеїнів). Дрібні органели (діаметром близько 20 нм). Відповідають за здійснення трансляції — синтезу поліпептидного ланцюга на матриці мРНК. Рибосома побудована із двох субодиниць — великої та малої, до складу кожної входить приблизно однакова за масою кількість білків та рРНК. Існує два основних типи рибосом — менші 70S, наявні у прокаріотичних клітинах, мітохондріях і пластидах, і дещо більші (80S рибосоми) цитоплазми еукаріот. В еукаріотичних клітинах виділяють дві основні популяції рибосом: вільні та пов'язані з ендоплазматичним ретикулумом (ЕПР). Ці дві групи не відрізняються за структурою, а лише синтезованими білками: вільні рибосоми синтезують цитоплазматичні білки, тоді як на шЕПР відбувається синтез мембранних і секреторних білків. Часто кілька рибосом рухаються одна за одною вздовж одного ланцюга мРНК, синтезуючи поліпептидні ланцюги, такі об'єднання рибосом називають полірибосомами або полісомами. Атомарна модель будови 50S рибосоми Haloarcula marismortui
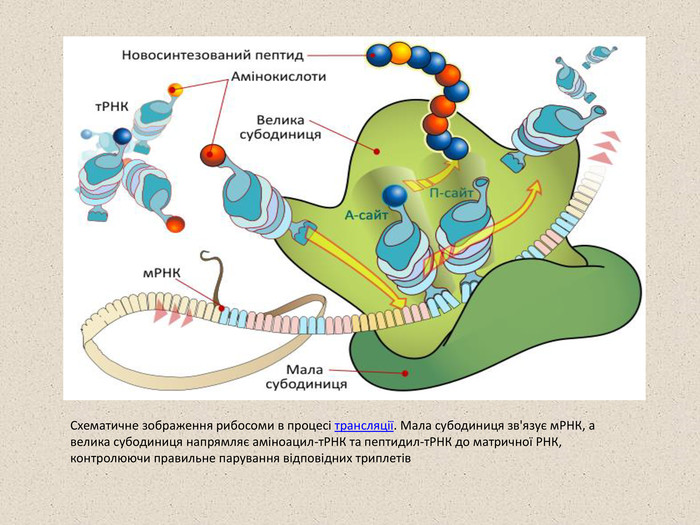
Схематичне зображення рибосоми в процесі трансляції. Мала субодиниця зв'язує мРНК, а велика субодиниця напрямляє аміноацил-тРНК та пептидил-тРНК до матричної РНК, контролюючи правильне парування відповідних триплетів
http://www.youtube.com/watch?v=8jFxVzspMTY&feature=fvwrel

БІОСИНТЕЗ БІЛКА http://www.youtube.com/watch?v=xIvexSkcorE&feature=related

ЕНДОМЕМБРАННА СИСТЕМА КЛІТИНИ Більшість мембран еукаріотичної клітини є частиною ендомембранної системи, функціями якої є здійснення кінцевих етапів біосинтезу більшості білків, та їх транспорт до відповідних органел або назовні клітини, метаболізм і транспорт ліпідів та детоксикація отруйних речовин. Всі мембрани цієї системи або безпосередньо переходять одна в одну, або пов'язані за допомогою маленьких мембранних мішечків — везикул, проте, не зважаючи, на такий зв'язок вони можуть суттєво відрізнятись за властивостями та функціями. До ендомембранної системи належать ендоплазматичний ретикулум (ЕПР) або ендоплазматична сітка, ядерна оболонка, апарат (комплекс) Гольджі, лізосоми, секреторні везикули та плазмалема.

ЕНДОПЛАЗМАТИЧНИЙ РЕТИКУЛУМ Ендоплазматичний ретикулум (ЕПР, від лат. reticulum — «сіточка») або ендоплазматична мережа — внутріклітинна органела еукаріотичних клітин, що представляє собою розгалужену систему з оточених мембраною сплющених порожнин, бульбашок і канальців. Клітина легень: у правому нижньому куті видно ядро, більшість клітини заповнено мембранами ендоплазматичного ретикулуму (ТЕМ)

Мембрани ЕПР зазвичай становлять більше половини загальної площі мембран клітини, вони утворюють сітку із трубочок та сплощених мішечків, котрі називають цистернами. Ці мембрани відділяють від цитоплазми окремий компартмент — просвіт ендоплазматичного ретикулуму, що займає приблизно 10% об'єму клітини і є неперервним із перинуклеарним простором. Виділяють два види ЕПР, що відрізняться за структурою та функціями: гладкий (агранулярний) ЕПР (гЕПР), на поверхні якого не розташовані рибосоми, та шорсткий або гранулярний ЕПР (шЕПР), який ними всіяний. Гладкий ендоплазматичний ретикулум бере участь у біосинтезі ліпідів (зокрема фосфоліпідів та стероїдних гормонів) та вуглеводів, а також у детоксикації отрут. Особливо багаті на гладкий ендоплазматичний ретикулум гепатоцити — клітини печінки, оскільки там інтенсивно відбувається метаболізм чужорідних речовин, зокрема фармацевтичних препаратів. Тривале вживання деяких препаратів, зокрема барбітуратів, стимулює збільшення кількості мембран гладкого ЕПР, через що збільшується і стійкість організму до дії не лише цих, а й інших медикаментів. Особливим типом гладкого ЕПР є саркоплазматичний ретикулум м'язових волокон, який накопичує у собі велику кількість іонів Ca+ і вивільняє їх у цитоплазму під час м'язового скорочення. Шорсткий ендоплазматичний ретикулум відрізняється від гладкого наявністю великої кількості рибосом на його поверхні. До головних функцій шЕПР належить здійснення кінцевих етапів біосинтезу секреторних білків, їх сортування та транспорт, а також утворення мембран клітини. Під час трансляції, що відбувається на мембранозв'язаних рибосомах, поліпептидний ланцюг транспортується у порожнину ЕПР через спеціальний білковий комплекс, там відбувається фолдинг білка — тобто утворення його просторової структури, а також, у багатьох випадках, модифікація, наприклад приєднання вуглеводних залишків. Після цього зрілі білки транспортуються за допомогою особливих везикул до місця призначення. Шорсткий ендоплазматичний ретикулум також бере участь у синтезі, модифікації і сортуванні мембранних білків та включенні у мембрани нових молекул фосфоліпідів.

КОМПЛЕКС ГОЛЬДЖІ Структуру відому зараз під назвою апарат Гольджі відкрив 1898 року Каміло Гольджі. Ця органела міститься майже в усіх еукаріотичних клітинах, особливо добре розвинена в тих, що виконують секреторну функцію. Комплекс Гольджі складається із великої кількості плоских мембранних мішечків — цистерн, ніби складених на стопку, і пов'язаної із ними системи пухирців — везикул Гольджі, що здійснюють транспорт між частинами апарату Гольджі, а також між апаратом Гольджі та іншими частинами клітини. Стопка цистерн апарату Гольджі або диктіосома характеризується полярністю: тобто дві її сторони відрізняються за структурою і функціями. Цис сторона зазвичай повернута в бік до ендоплазматичного ретикулуму: від ЕПР відшнуровуються везикули, які зливаються із цистернами цієї сторони, вивільняючи свій вміст в її просвіт. Поступово рухаючись у цистернах апарату Гольджі від цис до транс сторони молекули зазнають модифікації, наприклад у багатьох глікопротеїнів змінюються вуглеводні залишки. Окрім цього, комплекс Гольджі містить власні ферменти, що синтезують деякі речовини, наприклад у рослинних клітин, це пектини та інші компоненти клітинної стінки, відмінні від целюлози. Згодом модифіковані або новосинтезовані молекули потрапляють у мембранні пухирці, що відшнуровуються від транс сторони апарату Гольджі, і транспортуються до інших органел, або виводяться назовні клітини шляхом екзоцитозу. Ділянка лейкоцита: видно апарат Гольджі (ТЕМ)

ЛІЗОСОМИ Лізосома (від грецького 'lizis' - розчинення) - одномембранна органела сферичної форми, яка являє собою невелику мембранну везикулу, наповнену гідролітичними ферментами(протеази, ліпази, амілази, нуклеази), необхідних для контролювання внутрішньоклітинного розщеплення. Основна її функція - перетравлення відмерлих решток клітини. Була відкрита бельгійським цитологом Крістіаном де Дювом в 1949 році. Лізосоми наявні здебільшого, у тваринних клітинах. Оскільки гідролітичні ферменти найкраще працюють за низьких значень pH, у лізосомах підтримується кисле середовище. Білки лізосом синтезуються рибосомами на поверхні шорсткого ендоплазматичного ретикулуму, потім транспортуються до апарату Гольджі, де зазнають подальшої модифікації, після цього від транс сторони відшнуровуються первинні лізосоми, що містять ці білки. Первинні лізосоми можуть зливатись із фагосомами — везикулами утвореними внаслідок фагоцитозу, таким чином утворюються вторинні лізосоми, де відбувається розщеплення макромолекул до мономерів, які транспортуються у цитоплазму. Багато найпростіших живляться фагоцитуючи часточки їжі, їхні вторинні лізосоми називаються травними вакуолями. Деякі людські клітини також здатні до активного фагоцитозу, наприклад макрофаги та нейтрофіли. Лізосоми беруть участь в автофагії, під час цього процесу ушкоджені органели оточуються подвійною мембраною, з якою згодом зливається лізосома і перетравлює все, що було всередині, утворені при цьому мономери виходять у цитоплазму, і можуть використовуватись для побудови нових органел. Таким чином клітина постійно оновлюється.
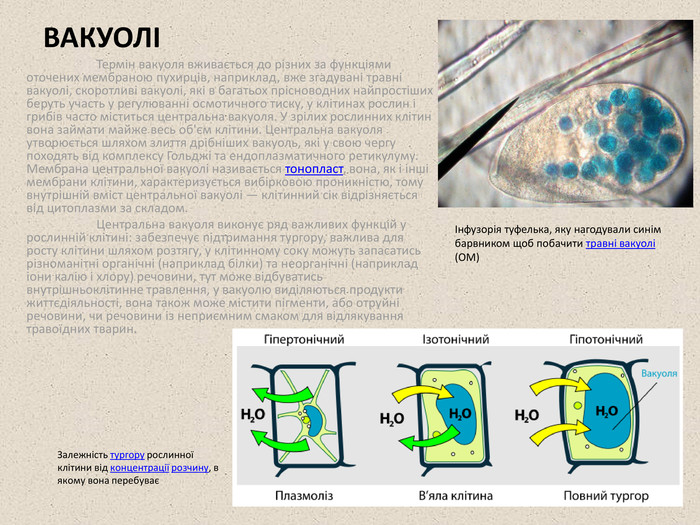
ВАКУОЛІ Термін вакуоля вживається до різних за функціями оточених мембраною пухирців, наприклад, вже згадувані травні вакуолі, скоротливі вакуолі, які в багатьох прісноводних найпростіших беруть участь у регулюванні осмотичного тиску, у клітинах рослин і грибів часто міститься центральна вакуоля. У зрілих рослинних клітин вона займати майже весь об'єм клітини. Центральна вакуоля утворюється шляхом злиття дрібніших вакуоль, які у свою чергу походять від комплексу Гольджі та ендоплазматичного ретикулуму. Мембрана центральної вакуолі називається тонопласт, вона, як і інші мембрани клітини, характеризується вибірковою проникністю, тому внутрішній вміст центральної вакуолі — клітинний сік відрізняється від цитоплазми за складом. Центральна вакуоля виконує ряд важливих функцій у рослинній клітині: забезпечує підтримання тургору, важлива для росту клітини шляхом розтягу, у клітинному соку можуть запасатись різноманітні органічні (наприклад білки) та неорганічні (наприклад іони калію і хлору) речовини, тут може відбуватись внутрішньоклітинне травлення, у вакуолю виділяються продукти життєдіяльності, вона також може містити пігменти, або отруйні речовини, чи речовини із неприємним смаком для відлякування травоїдних тварин. Інфузорія туфелька, яку нагодували синім барвником щоб побачити травні вакуолі (ОМ) Залежність тургору рослинної клітини від концентрації розчину, в якому вона перебуває

ПЕРОКСИСОМИ Пероксисоми — органели, присутні в усіх головних групах еукаріот, оточені однією мембраною, всередині містять ферменти, такі як каталаза та уратоксидаза, у такій великій кількості, що вони часто кристалізуються в центрі органели. До основних функцій пероксисом належить окиснення багатьох органічних речовин (зокрема β-окиснення жирних кислот, яке у тварин відбувається також і в мітохондріях, а в рослин та грибів — тільки у пероксисомах), знешкодження надлишку шкідливого для клітини пероксиду водню, метаболізм спиртів та амінів (наприклад 25% етилового спирту в печінці людини окиснюється саме в пероксисомах), здійснення гліоксалатного циклу у клітинах насіння рослин. Існують різні версії щодо походження нових пероксисом у клітині: вони можуть утворюватись шляхом поділу вже існуючих пероксисом і рости, транспортуючи білки і фосфоліпіди із цитоплазми, або з особливих везикул ендоплазматичного ретикулуму. Можливо, обидва описані процеси поєднуються в еукаріотичних клітинах. Пероксисома — органела, притаманна більшості еукаріотичних клітин. Носить назву гліоксисома в рослинах, глікосома в трипаносомах та тільця Вороніна в деяких видах грибів. Схематичне зображення пероксисоми
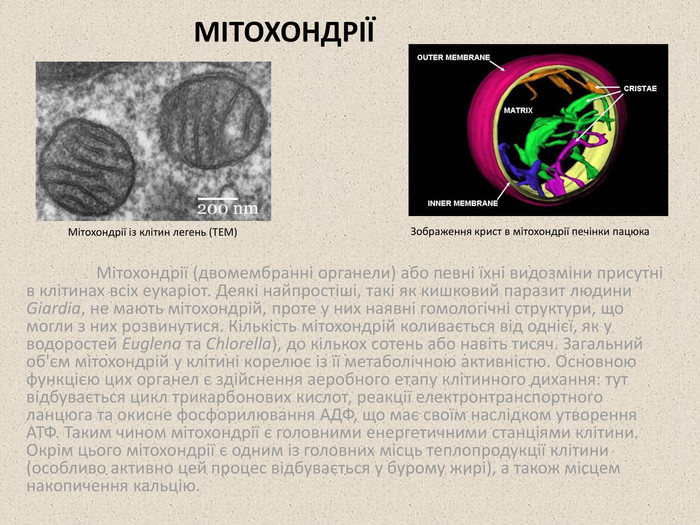
МІТОХОНДРІЇ Мітохондрії (двомембранні органели) або певні їхні видозміни присутні в клітинах всіх еукаріот. Деякі найпростіші, такі як кишковий паразит людини Giardia, не мають мітохондрій, проте у них наявні гомологічні структури, що могли з них розвинутися. Кількість мітохондрій коливається від однієї, як у водоростей Euglena та Chlorella), до кількох сотень або навіть тисяч. Загальний об'єм мітохондрій у клітині корелює із її метаболічною активністю. Основною функцією цих органел є здійснення аеробного етапу клітинного дихання: тут відбувається цикл трикарбонових кислот, реакції електронтранспортного ланцюга та окисне фосфорилювання АДФ, що має своїм наслідком утворення АТФ. Таким чином мітохондрії є головними енергетичними станціями клітини. Окрім цього мітохондрії є одним із головних місць теплопродукції клітини (особливо активно цей процес відбувається у бурому жирі), а також місцем накопичення кальцію. Мітохондрії із клітин легень (ТЕМ) Зображення крист в мітохондрії печінки пацюка
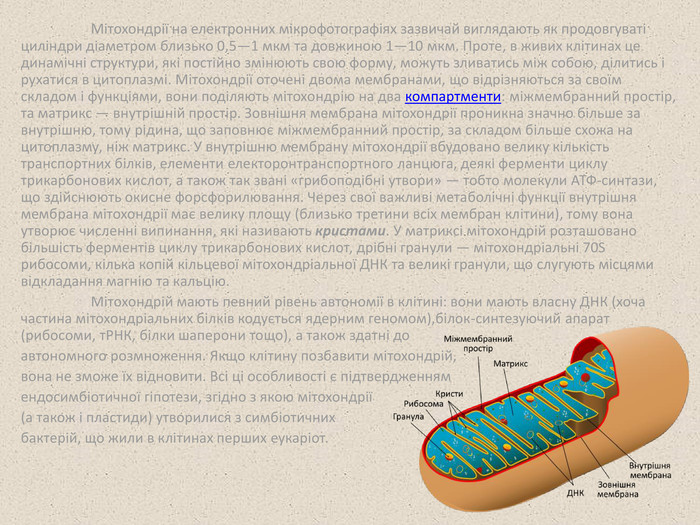
Мітохондрії на електронних мікрофотографіях зазвичай виглядають як продовгуваті циліндри діаметром близько 0,5—1 мкм та довжиною 1—10 мкм. Проте, в живих клітинах це динамічні структури, які постійно змінюють свою форму, можуть зливатись між собою, ділитись і рухатися в цитоплазмі. Мітохондрії оточені двома мембранами, що відрізняються за своїм складом і функціями, вони поділяють мітохондрію на два компартменти: міжмембранний простір, та матрикс — внутрішній простір. Зовнішня мембрана мітохондрії проникна значно більше за внутрішню, тому рідина, що заповнює міжмембранний простір, за складом більше схожа на цитоплазму, ніж матрикс. У внутрішню мембрану мітохондрії вбудовано велику кількість транспортних білків, елементи електоронтранспортного ланцюга, деякі ферменти циклу трикарбонових кислот, а також так звані «грибоподібні утвори» — тобто молекули АТФ-синтази, що здійснюють окисне форсфорилювання. Через свої важливі метаболічні функції внутрішня мембрана мітохондрії має велику площу (близько третини всіх мембран клітини), тому вона утворює численні випинання, які називають кристами. У матриксі мітохондрій розташовано більшість ферментів циклу трикарбонових кислот, дрібні гранули — мітохондріальні 70S рибосоми, кілька копій кільцевої мітохондріальної ДНК та великі гранули, що слугують місцями відкладання магнію та кальцію. Мітохондрій мають певний рівень автономії в клітині: вони мають власну ДНК (хоча частина мітохондріальних білків кодується ядерним геномом),білок-синтезуючий апарат (рибосоми, тРНК, білки шаперони тощо), а також здатні до автономного розмноження. Якщо клітину позбавити мітохондрій, вона не зможе їх відновити. Всі ці особливості є підтвердженням ендосимбіотичної гіпотези, згідно з якою мітохондрії (а також і пластиди) утворилися з симбіотичних бактерій, що жили в клітинах перших еукаріот.
http://www.youtube.com/watch?v=aSSpHC82m_M&feature=related

ПЛАСТИДИ Пластиди наявні в усіх живих рослинних клітинах. Ці органели між собою поєднує те, що їх вкрито двома мембранами і в одного організму є кілька однакових копій ДНК. Всі пластиди утворюються з пропластид меристемних клітин рослин. Пропластиди диференціюються залежно від потреб клітини: хлоропласти — зелені пластиди, що містять хлорофіл і здійснюють фотосинтез; етіопласти утворюються в темряві, вони містять жовтий пігмент — попередник хлорофілу, на світлі можуть швидко перетворюватись на хлоропласти; лейкопласти — утворюються у нефотосинтетичних клітинах рослин, є місцем запасання органічних речовин, зокрема в амілопластах накопичуються вуглеводи (крохмаль), в елайопластах — жири, у протеїнопластах — білки; хромопласти — червоні, жовті або оранжеві пластиди, накопичують пігменти, зокрема каротиноїди. Окрім фотосинтезу та накопичення різних речовин, у пластидах також відбуваються процеси синтезу пуринів та піримідинів, жирних кислот та деяких амінокислот тощо. Пластиди, як і мітохондрії, є порівняно автономними органелами клітини. Вважається, що вони можуть походити від симбіотичних ціанобактерій. Основні типи пластид в рослинній клітині
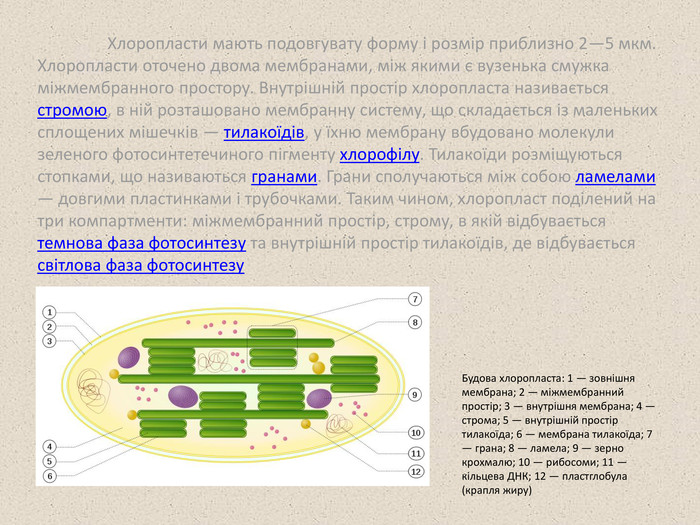
Хлоропласти мають подовгувату форму і розмір приблизно 2—5 мкм. Хлоропласти оточено двома мембранами, між якими є вузенька смужка міжмембранного простору. Внутрішній простір хлоропласта називається стромою, в ній розташовано мембранну систему, що складається із маленьких сплощених мішечків — тилакоїдів, у їхню мембрану вбудовано молекули зеленого фотосинтетечиного пігменту хлорофілу. Тилакоїди розміщуються стопками, що називаються гранами. Грани сполучаються між собою ламелами — довгими пластинками і трубочками. Таким чином, хлоропласт поділений на три компартменти: міжмембранний простір, строму, в якій відбувається темнова фаза фотосинтезу та внутрішній простір тилакоїдів, де відбувається світлова фаза фотосинтезу Будова хлоропласта: 1 — зовнішня мембрана; 2 — міжмембранний простір; 3 — внутрішня мембрана; 4 — строма; 5 — внутрішній простір тилакоїда; 6 — мембрана тилакоїда; 7 — грана; 8 — ламела; 9 — зерно крохмалю; 10 — рибосоми; 11 — кільцева ДНК; 12 — пластглобула (крапля жиру)
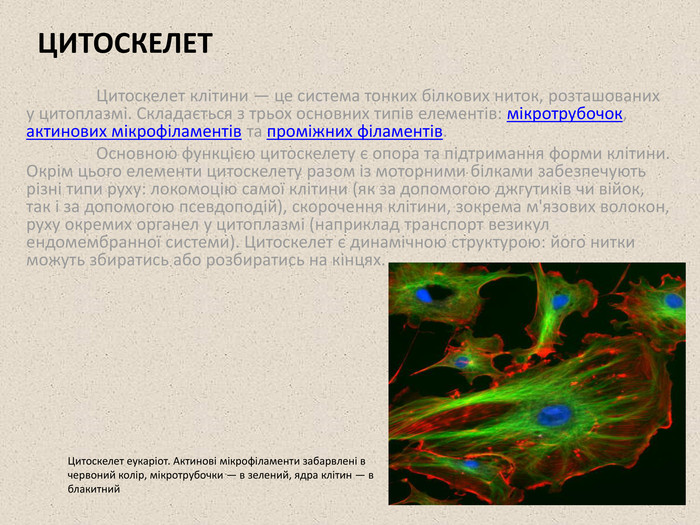
ЦИТОСКЕЛЕТ Цитоскелет клітини — це система тонких білкових ниток, розташованих у цитоплазмі. Складається з трьох основних типів елементів: мікротрубочок, актинових мікрофіламентів та проміжних філаментів. Основною функцією цитоскелету є опора та підтримання форми клітини. Окрім цього елементи цитоскелету разом із моторними білками забезпечують різні типи руху: локомоцію самої клітини (як за допомогою джгутиків чи війок, так і за допомогою псевдоподій), скорочення клітини, зокрема м'язових волокон, руху окремих органел у цитоплазмі (наприклад транспорт везикул ендомембранної системи). Цитоскелет є динамічною структурою: його нитки можуть збиратись або розбиратись на кінцях. Цитоскелет еукаріот. Актинові мікрофіламенти забарвлені в червоний колір, мікротрубочки — в зелений, ядра клітин — в блакитний

Мікротрубочки, клітинний центр та джгутики Мікротрубочки — це порожнисті циліндри діаметром 25 нм і довжиною 0,2—25 мкм, що складаються зі спірально розташованих димерів білка тубуліну. Вони можуть рости або зменшуватися шляхом полімеризації або дисоціації тубуліну на одному з кінців. Мікротрубочки беруть участь у підтриманні форми клітини, зокрема запобігають її стисканню, у внутрішньоклітинному транспорті, а також забезпечують розходження хроматид (або хромосом) під час клітинного поділу. У тваринній клітині мікротрубочки «ростуть» із клітинного центру (центросоми), розташованого поблизу ядра, він виконує функцію центру організації мікротрубочок (ЦОМ). На цій ділянці також є два короткі порожнисті циліндри (довжина 30—50 мкм, діаметр 20 мкм) — центріолі вони побудовані із дев'яти триплетів мікротрубочок, розміщених колом. Перед клітинним поділом центріолі подвоюються, кожна пара розходиться до одного з полюсів клітини, де вони стають центрами організації для мікротрубочок веретена поділу. Клітинний центр і центріолі виявлено тільки у тваринних клітинах, у рослин та грибів їх функції мають виконувати інші структури. Мікротрубочки також є основними структурними елементами джгутиків та війок — органел руху, наявних переважно у тваринних клітин. Джгутики та війки ідентичні за будовою, але відрізняються довжиною, кількістю на одну клітину та характером руху. Обидва типи органел складаються із двох основних частин: базального тіла, розташованого всередині клітини та аксонеми — довгої нитки, вкритої плазматичною мембраною. Базальне тіло схоже за структурою до центріолі — складається із дев'яти триплетів мікротрубочок. Усередині аксонеми також розташовано мікротрубочки, але іншим чином: дев'ять пар утворюють циліндр, усередині якого розміщена ще одна пара (принцип розміщення «9+2»). У русі джгутиків та війок беруть участь моторні білки динеїни. Переріз через аксонеми джгутиків хламідомонади (Chlamydomonas reinhardtii) (ТЕМ)
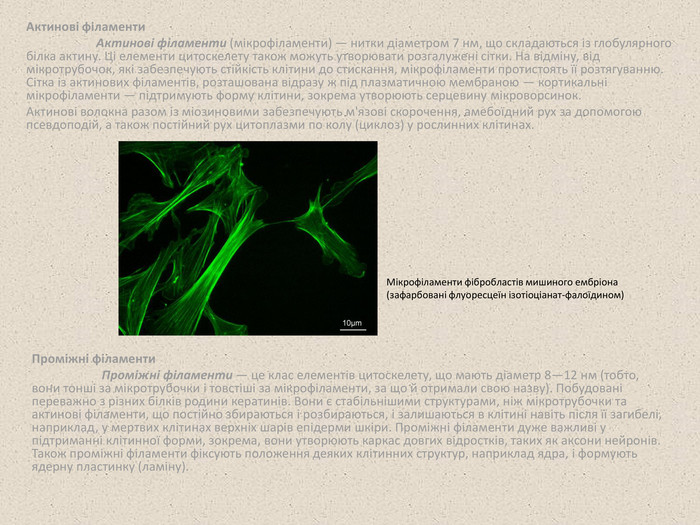
Актинові філаменти Актинові філаменти (мікрофіламенти) — нитки діаметром 7 нм, що складаються із глобулярного білка актину. Ці елементи цитоскелету також можуть утворювати розгалужені сітки. На відміну, від мікротрубочок, які забезпечують стійкість клітини до стискання, мікрофіламенти протистоять її розтягуванню. Сітка із актинових філаментів, розташована відразу ж під плазматичною мембраною — кортикальні мікрофіламенти — підтримують форму клітини, зокрема утворюють серцевину мікроворсинок. Актинові волокна разом із міозиновими забезпечують м'язові скорочення, амебоїдний рух за допомогою псевдоподій, а також постійний рух цитоплазми по колу (циклоз) у рослинних клітинах. Проміжні філаменти Проміжні філаменти — це клас елементів цитоскелету, що мають діаметр 8—12 нм (тобто, вони тонші за мікротрубочки і товстіші за мікрофіламенти, за що й отримали свою назву). Побудовані переважно з різних білків родини кератинів. Вони є стабільнішими структурами, ніж мікротрубочки та актинові філаменти, що постійно збираються і розбираються, і залишаються в клітині навіть після її загибелі, наприклад, у мертвих клітинах верхніх шарів епідерми шкіри. Проміжні філаменти дуже важливі у підтриманні клітинної форми, зокрема, вони утворюють каркас довгих відростків, таких як аксони нейронів. Також проміжні філаменти фіксують положення деяких клітинних структур, наприклад ядра, і формують ядерну пластинку (ламіну). Мікрофіламенти фібробластів мишиного ембріона (зафарбовані флуоресцеїн ізотіоціанат-фалоїдином)
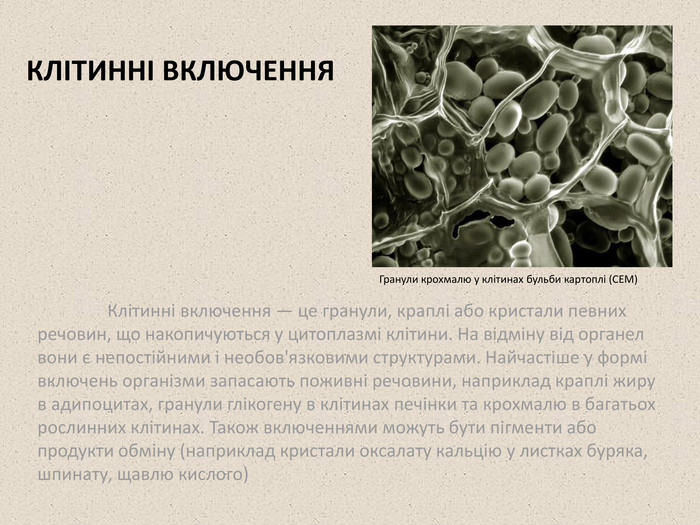
КЛІТИННІ ВКЛЮЧЕННЯ Клітинні включення — це гранули, краплі або кристали певних речовин, що накопичуються у цитоплазмі клітини. На відміну від органел вони є непостійними і необов'язковими структурами. Найчастіше у формі включень організми запасають поживні речовини, наприклад краплі жиру в адипоцитах, гранули глікогену в клітинах печінки та крохмалю в багатьох рослинних клітинах. Також включеннями можуть бути пігменти або продукти обміну (наприклад кристали оксалату кальцію у листках буряка, шпинату, щавлю кислого) Гранули крохмалю у клітинах бульби картоплі (СЕМ)

КЛІТИННА СТІНКА Клітинна стінка — це надмембранна структура, наявна у клітин рослин, грибів (а також у прокаріот), але відсутня у тварин. Вона потрібна для підтримання форми, захисту клітини та запобігання надмірного надходження до неї води. У грибів клітинна стінка складається в основному з хітину, а в рослин — із фібрил целюлози та геміцелюлоз, занурених у матрикс із пектинів. Молода рослинна клітина утворює тонку гнучку первинну клітинну стінку (товщиною близько 0,1 мкм). Між клітинними стінками двох сусідніх клітин розміщується серединна пластинка, що складається в основному із пектинів, які «склеюють» клітини між собою. Після того як рослинна клітина перестає рости, вона укріплює свою клітинну стінку, відкладаючи додаткові шари целюлози. У певних тканинах (наприклад провідних та опорних) клітини утворюють досить товсту вторинну клітинну стінку, що може складатись з інших речовин — наприклад лігніну в деревині або суберину у корку. Структура клітинної стінки рослинної клітини

УЗАГАЛЬНЕННЯ ЗНАНЬ http://www.youtube.com/watch?v=QXqhswX5rsc http://www.youtube.com/watch?v=-D74CNJQmkc


про публікацію авторської розробки
Додати розробку