Листок як орган фотосинтезу


« Листок як орган фотосинтезу»

Листок-орган фотосинтезу. У процесі еволюції рослин сформувався спеціалізований орган фотосинтезу – листок. У середньому листки поглинають 80–85 % енергії фотосинтетично активної радіації (ФАР), що є видимою частиною спектру електромагнітного випромінювання з довжиною хвилі 400–700 нм і 25 % енергії інфрачервоних променів, що становить близько 55 % енергії загальної радіації. Рис. 1 Рослина 2

Листок відбиває 10 % ФАР і 45 % інфрачервоних променів і пропускає відповідно 5 % і 30 %. На фотосинтез використається 1,5–2 % поглиненої синтетично активної радіації, інша поглинена енергія витрачається переважно на випаровування води – транспірацію (95–98 %), можливий також і теплообмін з атмосферою. Рис.2 Листок берези під мікроскопом3

Хлорофіли та каротиноїди Листкова поверхня досягає значних розмірів і перевершує площу ґрунту, що займає рослина. Для характеристики розмірів фотосинтетичного апарата використовують індекс листкової поверхні (L) – площа листя (м2 ), що припадає на 1 м2 ґрунту. Для сільськогосподарських рослин помірної зони середні значення листкового індексу варіює у межах 3–5, у південних широтах з вологим кліматом – до 8–10. Наприклад, поверхня листя 1 га посівів зернових культур у фазі цвітіння досягає 20–40 тис. м2 , у низькорослих яблунь під час вегетації площа листя становить 25–30 тис. м2 /га. Фотосинтез обумовлюється вибірковим поглинанням пігментами світла у видимій частині сонячного спектра. Частка фотосинтетичних пігментів становить 10–15 % сухої маси хлоропластів. За хімічним походженням фотосинтетичні пігменти поділяються на дві групи – хлорофіли та каротиноїди.4

Що таке каротиноїди?КАРОТИНОЇДИ (лат. carotinoida < лат. carota — морква) — жовті, жовтогарячі або червоні пігменти (циклічні чи ациклічні ізопреноїди), синтезовані бактеріями, грибами і вищими рослинами. Тварини зазвичай не утворюють К., але використовують їх для синтезу вітаміну А. До К. належать поширені в рослинах каротини і ксантофіли; лікопін (С40 Н56) — у плодах томатів, шипшини, пасльону; зеаксантин (С40 Н56 О2) — у насінні кукурудзи; віолаксантин і флавоксантин — у плодах гарбуза; Рис. 3 Овочі 5

Хлорофіли. Хлоропласти вищих рослин містять хлорофіл а й хлорофіл b. Вони були ідентифіковані російським вченим М. С. Цвєтом (1906 р.) за допомогою розробленого ним методу хроматографії. Структурна формула хлорофілу, запропонована Г. Фішером (1939 р.), одержала остаточне підтвердження в 1960 р. у результаті двох незалежно проведених робіт у США і ФРН зі штучного синтезу хлорофілу а. Рис. 4 Листок під мікроскопом 6

Фотосинтезуючі зелені бактеріїХлорофіл – складний ефір дикарбонової кислоти хлорофіліна, у якої одна карбоксильна група етерифікована залишком метилового спирту (СН3 ОН), а інша – залишком одноатомного неорганічного спирту фітолу (С20 Н39 ОН). У всіх вищих рослин, водоростей і ціанобактерій міститься хлорофіл a, хлорофіл b є у вищих рослин і зелених водоростей. Хлорофіл с, позбавлений фітолу, міститься в бурих і діатомових водоростях, хлорофіл d – у червоних водоростях. Рис. 5 Модель молекули хлорофілу 7

Структура циклопентанового кільця. У хлорофіла а чотири пірольних кільця з’єднані між собою метиновими містками (= СН-), утворюючи порфіринове кільце. Крім того, атоми азоту пірольних кілець пов’язані з атомом магнію. З порфіриновим ядром пов’язане циклопентанове кільце, утворене залишком кетопропіонової кислоти й містить активні карбонільну (С = О) і метильовану карбоксильну (О = С-О-СН3) групи. Структура, що складається з порфіринового ядра та циклопентанового кільця, називається форбіном. Рис. 6 Модель циклопениану 8

Хімічні властивостіЯкщо атом магнію заміщений протоном, то таке з’єднання називається феофітин. Активність хлорофілів, також як й інших пігментів, обумовлена наявністю великої кількості подвійних зв’язків з делокалізованими електронами. Хлорофіли добре розчинні в органічних розчинниках (етиловому ефірі, бензолі, хлороформі, ацетоні, етиловому спирті) і нерозчинні у воді. Хлорофіли мають максимуми поглинання світла в червоній і синій частинах спектра. Рис. 7 Структурна формула феофітину9
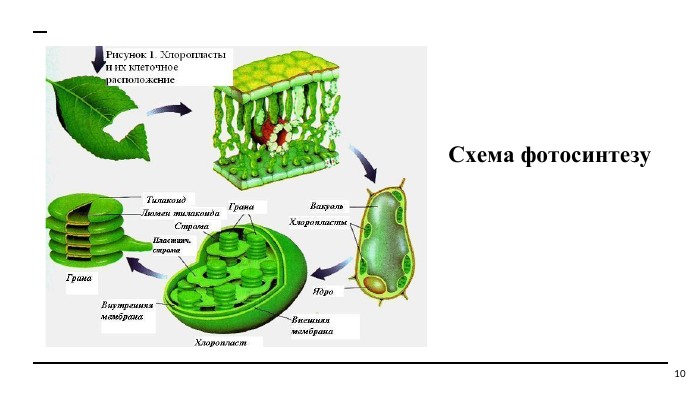
Схема фотосинтезу 10

Фотосинтез Фотосинтез характеризується швидкістю або інтенсивністю. Інтенсивність фотосинтезу вимірюється кількістю міліграмів СО2, який поглинений одним квадратним дециметром листової поверхні за 1 годину. Для її вимірювання визначають зменшення кількості вуглекислого газу в замкнутій камері, у якій розміщено листок. Одночасно разом із визначенням інтенсивності фотосинтезу визначають у темноті інтенсивність дихання цих клітин. Кількість СО2, що вивільняється під час дихання, додають до його кількості, що поглинена при освітленні. При цьому вважають, що інтенсивність дихання клітин на світлі та в темноті однакова. Однак, ці поправки не можуть точно оцінити дійсний фотосинтез, оскільки при затінені листка виключається не тільки фотосинтез, але й гліколатний цикл, а також тому, що дихання в дійсності трохи залежить від світла.11

Квантова витрата фотосинтезу. Величина фотосинтетичного коефіцієнта залежить від умов освітлення та мінерального живлення. Освітлення рослин синім світлом і гарне постачання азоту збільшує синтез білків, отже, фотосинтетичний коефіцієнт збільшується. Для характеристики фотосинтезу використовують такі показники: квантова витрата, квантовий вихід і асиміляційне число. Квантова витрата фотосинтезу – це співвідношення числа поглинених квантів світла до числа асимільованих молекул двоокису вуглецю12
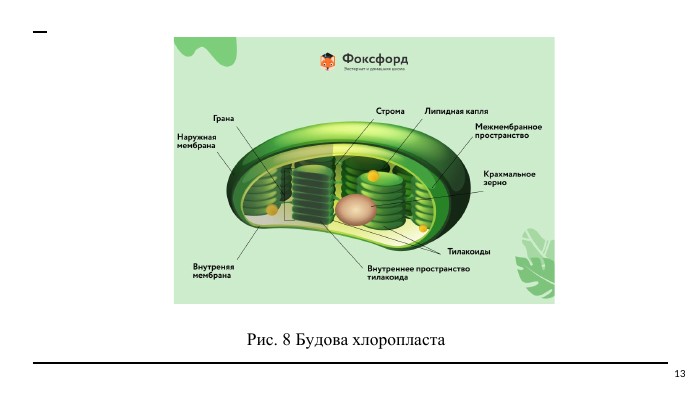
Рис. 8 Будова хлоропласта 13

Асиміляційне число Обернена величина отримала назву квантового виходу. Асиміляційне число – кількість асимільованої листом за 1 годину вуглекислоти, що припадає на одиницю хлорофілу, що міститься в листку. Кількість грамів сухої речовини, яка накопичена в рослині за добу в перерахунку на 1 м2 листкової поверхні, характеризує чисту продуктивність фотосинтезу (ЧПФ). Рис . 9 Будова СО214

Накопичення органічної маси. У процесі фотосинтезу наземні рослини утворюють до 170 млрд тон, а рослини світового океану – до 70 млрд тон біомаси в рік у перерахунку на суху речовину, що використовується гетеротрофними організмами. Забезпечення сталості вмісту СО2 в повітрі. Зв’язування СО2 у процесі фотосинтезу значною мірою компенсує його виділення внаслідок інших процесів (дихання, бродіння, діяльність вулканів, виробнича діяльність людства). 15

Основне про листок. Листок у процесі формування проходить чотири фази: утворення примордію; формування осі листка (черешка); закладання пластинки листка за рахунок бокової меристеми; ріст пластинки шляхом розтягування. Кожен листковий примордій утворюється як горбик в периферичній меристемі апекса пагона внаслідок локального поділу клітин (площина поділу паралельна поверхні апекса). Примордій бічної (пазушної) бруньки з’являється дещо пізніше. Згодом у ньому формується апікальна меристема, гомологічна до апексу головного пагона. Зачатки листків впливають на тканини, що розміщуються нижче, індукуючи диференціацію провідних пучків. 16
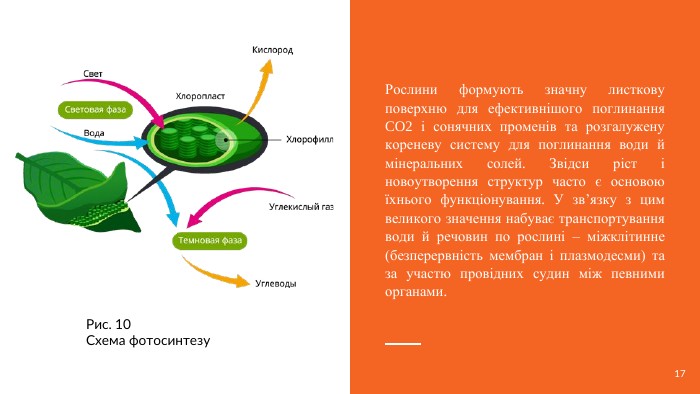
Рослини формують значну листкову поверхню для ефективнішого поглинання CO2 і сонячних променів та розгалужену кореневу систему для поглинання води й мінеральних солей. Звідси ріст і новоутворення структур часто є основою їхнього функціонування. У зв’язку з цим великого значення набуває транспортування води й речовин по рослині – міжклітинне (безперервність мембран і плазмодесми) та за участю провідних судин між певними органами. Рис. 10 Схема фотосинтезу17

Функціональні властивості рослин. Транспортні системи об’єднують органи й тканини в єдиний організм. Вони мають важливе значення для регуляції функції росту й розвитку рослин. Специфіка метаболізму в окремих органах і обмін речовин створюють основу взаємодії органів, обумовлюють цілісність рослинного організму. Ріст є основою формування всіх функцій, і кожна функція, у свою чергу, зазнає певних змін в онтогенезі рослин. Однією з важливих функціональних властивостей рослин є здатність адаптуватися до зміни умов навколишнього середовища, що забезпечує стійкість рослин до абіотичних і біотичних стресів. Фізіологія рослин вивчає перетворення речовини, енергії та форми у рослин, а також перетворення інформації. 18

Висновок. Рослини мають велику листкову поверхню. Вона полегшує поглинання CО2, уловлювання світла та створює поверхню випаровування. Вода випаровується з поверхні листя через клітинні стінки епідермальних клітин і покривні шари (кутикулярна транспірація) і через продихи (продихова транспірація). Унаслідок втрати води під час транспірації в клітинах листя зростає всисна сила. Це призводить до посилення поглинання клітинами листка води з судин ксилеми й пересуванню води по ксилемі з коренів в листя. Отже, верхній кінцевий двигун, який бере участь у транспортуванні води вгору по рослині, обумовлений транспирацією листя.19

Список використаної літератури. Брайон О. В. Анатомія рослин / О. В. Брайон. – Київ : Вища школа, 1992. – 272 с. Власенко М. Ю. Фізіологія рослин / М. Ю. Власенко, Л. Д. Вельямінова-Зернова, В. В. Мацкевич. – Біла Церква : Білоцерківський державний аграрний університет, 2006. – 504 с. Грицаєнко З. М. Методи біологічних та агрохімічних досліджень рослин і ґрунтів / З. М. Грицаєнко, А. О. Грицаєнко, В. П. Карпенко. – Київ : ЗАТ «Нічлава», 2003. – 316 с. Злобін Ю. А. Курс фізіології і біохімії рослин / Ю. А. Злобін. – Суми : Університетська книга, 2004. – 464 с. Кузнецов В. В. Физиология растений : учебник для вузов / В. В. Кузнецов, Г. А. Дмитриева. – М. : Высш. шк., 2005. – 736 с. 20

Дякую за увагу!21


про публікацію авторської розробки
Додати розробку